10. Sistema Nervioso
El sistema nervioso permite al organismo reaccionar frente a los continuos cambios que se producen en el medio ambiente y en el medio interno. Además, controla e integra las diversas actividades del organismo, como la circulación y la respiración. Con fines descriptivos, el sistema nervioso se divide:
- Estructuralmente, en sistema nervioso central (SNC), compuesto por el encéfalo y la médula espinal, y sistema nervioso periférico (SNP), es decir, el resto del sistema nervioso que no pertenece al SNC.
- Funcionalmente, en sistema nervioso somático (SNS) y sistema nervioso autónomo (SNA).
El tejido nervioso se compone de dos tipos principales de células: neuronas (células nerviosas) y neuroglia (células de la glia), que sirven de soporte a las neuronas.
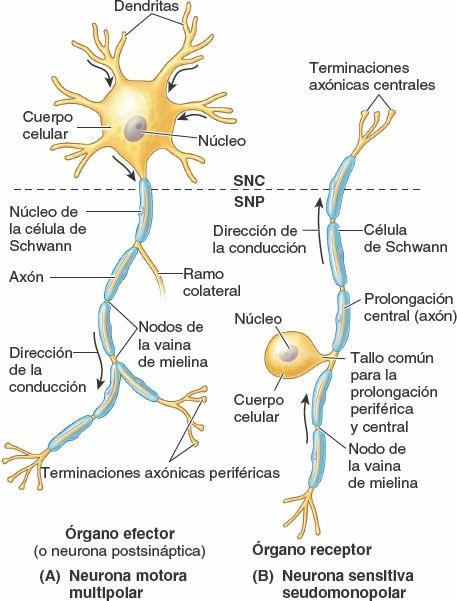
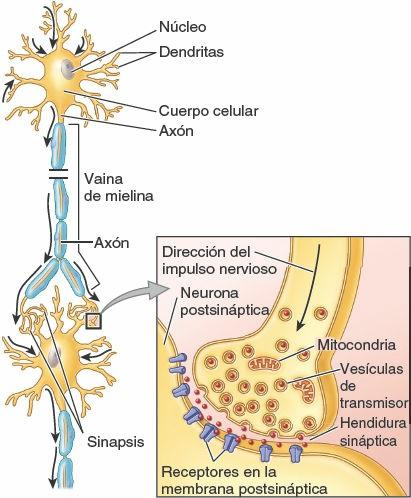
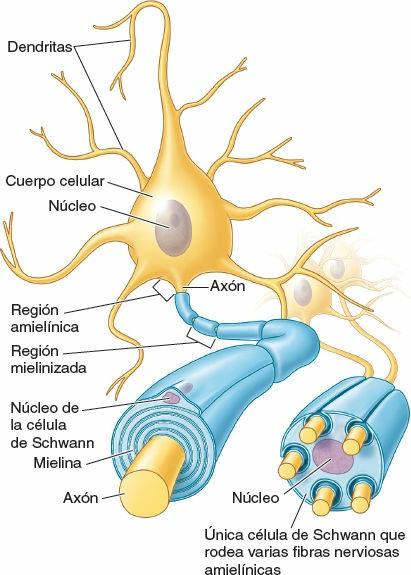
Las neuronas son las unidades estructurales y funcionales del sistema nervioso, especializadas para una rápida comunicación (figs. 1-28 y 1-29). Una neurona se compone del cuerpo celular, con prolongaciones (extensiones) denominadas dendritas y un axón, que llevan los impulsos hacia y desde el cuerpo celular, respectivamente. La mielina, capas de sustancias lipídicas y proteicas, forma una vaina de mielina en torno a algunos axones, lo que aumenta considerablemente la velocidad de conducción de los impulsos. Dos tipos de neuronas constituyen la mayoría de las existentes en el SNC (y en particular en el SNP) (fig. 1-28):
- Las neuronas motoras multipolares poseen dos o más dendritas y un solo axón, que puede tener uno o más ramos colaterales. Son el tipo más habitual de neurona en el sistema nervioso (SNC y SNP). Todas las neuronas motoras que controlan los músculos esqueléticos y las que componen el SNA son neuronas multipolares.
- Las neuronas sensitivas seudomonopolares poseen una corta prolongación, aparentemente única (pero en realidad doble), que se extiende desde el cuerpo celular. Esta prolongación común se separa en una prolongación periférica, que conduce los impulsos desde el órgano receptor (ej. sensores del tacto, el dolor o la temperatura en la piel) hacia el cuerpo celular, y una prolongación central, que continúa desde el cuerpo celular hasta el SNC. Los cuerpos celulares de las neuronas seudomonopolares están localizados fuera del SNC, en los ganglios sensitivos; por lo tanto, forman parte del SNP. Las neuronas comunican unas con otras en las sinapsis, o puntos de contacto interneuronales (fig. 1-29). La comunicación se produce por medio de neurotransmisores, sustancias químicas liberadas o secretadas por una neurona que pueden excitar o inhibir a otra, lo que continúa o interrumpe la conexión de los impulsos o la respuesta a ellos.
Las células de la neuroglia (células gliales o glia) son aproximadamente cinco veces más abundantes que las neuronas. Son células no neuronales ni excitables que constituyen un componente principal del tejido nervioso, con las funciones de apoyar, aislar o nutrir a las neuronas. En el SNC, la neuroglia incluye la oligodendroglia, los astrocitos, las células ependimarias y la microglia (pequeñas células gliales). En el SNP, la neuroglia comprende las células satélites en torno a las neuronas de los ganglios sensitivos de los vervios espinales (raíz posterior) y los ganglios del SNA, y las células de Schwann (neurilema) (figs. 1-28 y 1-29).
Sistema nervioso central
El sistema nervioso central (SNC) se compone del encéfalo y la médula espinal (fig. 1-30). Sus funciones principales consisten en integrar y coordinar las señales nerviosas de entrada y salida, y llevar a cabo las funciones mentales superiores, como el pensamiento y el aprendizaje.
Un núcleo es un acúmulo de cuerpos de neuronas en el SNC. Un haz de fibras nerviosas (axones) dentro del SNC que conecta núcleos de la corteza cerebral, cercanos o distantes, es un tracto. El encéfalo y la médula espinal se componen de sustancia gris y sustancia blanca. Los cuerpos de las neuronas constituyen la sustancia gris; los sistemas de tractos de fibras de interconexión forman la sustancia blanca (fig. 1-31). En los cortes transversales de la médula espinal, la sustancia gris presenta una forma parecida a una H, envuelta por una matriz de sustancia blanca. Los puntales (soportes) de la H son los cuernos (astas); por lo tanto, hay cuernos grises posteriores (dorsales) y anteriores (ventrales), derechos e izquierdos.
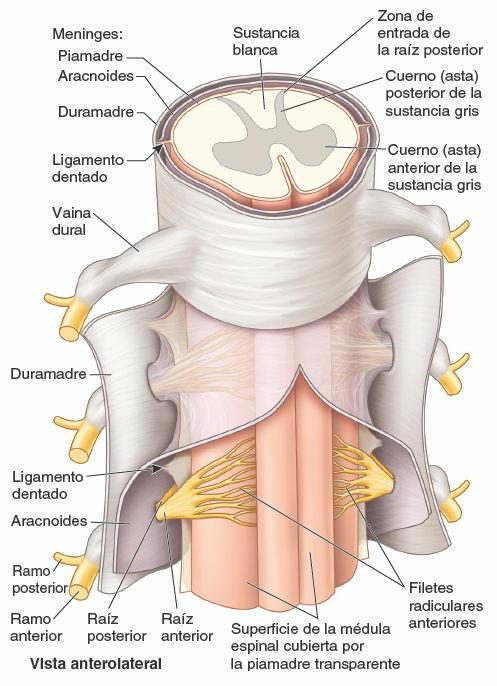
Tres capas membranosas (piamadre, aracnoides y duramadre) constituyen conjuntamente las meninges. Las meninges y el líquido cerebroespinal (LCE) rodean al SNC y lo protegen. El encéfalo y la médula espinal se hallan íntimamente recubiertos en su superficie externa por la capa meníngea más interna, una fina cubierta transparente, la piamadre. El LCE se halla entre la piamadre y la aracnoides. Por fuera de la piamadre y la aracnoides se encuentra la duramadre, firme y gruesa.
La duramadre del encéfalo está íntimamente relacionada con la cara interna de los huesos del neurocráneo circundante; la duramadre de la médula espinal está separada de los huesos de la columna vertebral por el espacio epidural, lleno de tejido adiposo.
Sistema nervioso periférico
El sistema nervioso periférico (SNP) se compone de fibras nerviosas y cuerpos celulares, situados fuera del SNC, que conducen los impulsos hacia o desde este (fig. 1-30). El SNP está organizado en nervios que conectan el SNC con las estructuras periféricas.
Una fibra nerviosa consta de un axón, su neurilema (del griego neuron, nervio + lemma, cáscara) y el tejido conectivo endoneural circundante (fig. 1-32). El neurilema está formado por la membrana de las células de Schwann, que rodea inmediatamente el axón y lo separa de otros axones. En el SNP, el neurilema puede adoptar dos formas, lo que crea dos clases de fibras nerviosas:
- El neurilema de las fibras nerviosas mielinizadas consiste en células de Schwann específicas para un determinado axón, organizadas en una serie continua de células envolventes formadoras de mielina.
- El neurilema de las fibras nerviosas amielínicas está formado por células de Schwann que no componen una serie aparente; hay múltiples axones incluidos por separado en el citoplasma de cada célula. Estas células de Schwann no producen mielina. La mayoría de las fibras de los nervios cutáneos (los nervios que confieren sensibilidad a la piel) son amielínicas.
Un nervio consta de:
- Un haz de fibras nerviosas situadas fuera del SNC (o un «acúmulo de fibras amontonadas», o fascículo, en los nervios de mayor tamaño).
- Las coberturas de tejido conectivo que rodean y unen las fibras nerviosas y los fascículos.
- Los vasos sanguíneos (vasa nervorum) que nutren las fibras nerviosas y sus cubiertas (fig. 1-33).
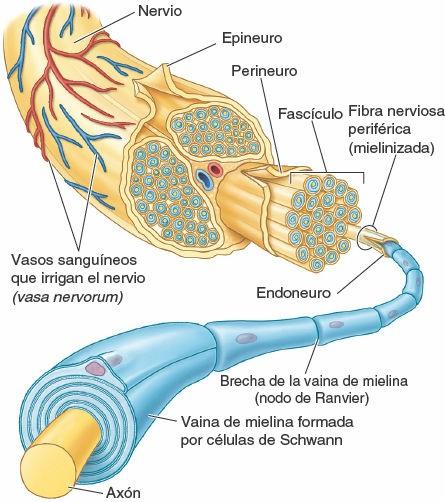
Los nervios son bastante fuertes y resistentes, porque sus fibras reciben soporte y protección de tres coberturas de tejido conectivo:
- Endoneuro, tejido conectivo fino que rodea íntimamente las células del neurilema y los axones.
- Perineuro, una capa de tejido conectivo denso que engloba un fascículo de fibras nerviosas y constituye una eficaz barrera contra las sustancias extrañas que pudieran penetrar en dichas fibras.
- Epineuro, una lámina gruesa de tejido conectivo que rodea y engloba un haz de fascículos, y forma la cobertura más externa del nervio; incluye tejido adiposo, vasos sanguíneos y linfáticos.
Los nervios están organizados de un modo muy semejante a un cable telefónico.
Los axones son como los hilos individuales aislados por el neurilema y el endoneuro; los hilos aislados son agrupados por el perineuro, y los grupos están rodeados por el epineuro, que forma la envoltura más externa del cable (fig. 1-33). Es importante distinguir entre fibras nerviosas y nervios, que a veces se representan igual en las figuras.
Un conjunto de cuerpos neuronales fuera del SNC constituye un ganglio. Existen ganglios motores (autónomos) y ganglios sensitivos.
Tipos de nervios
El SNP se continúa anatómica y operativamente con el SNC (fig. 1-30). Sus fibras aferentes (sensitivas) llevan los impulsos nerviosos al SNC, procedentes de los órganos de los sentidos (ej. el ojo) y de los receptores sensitivos de diversas partes del cuerpo (ej. en la piel). Sus fibras eferentes (motoras) conducen los impulsos nerviosos desde el SNC a los órganos efectores (músculos y glándulas).
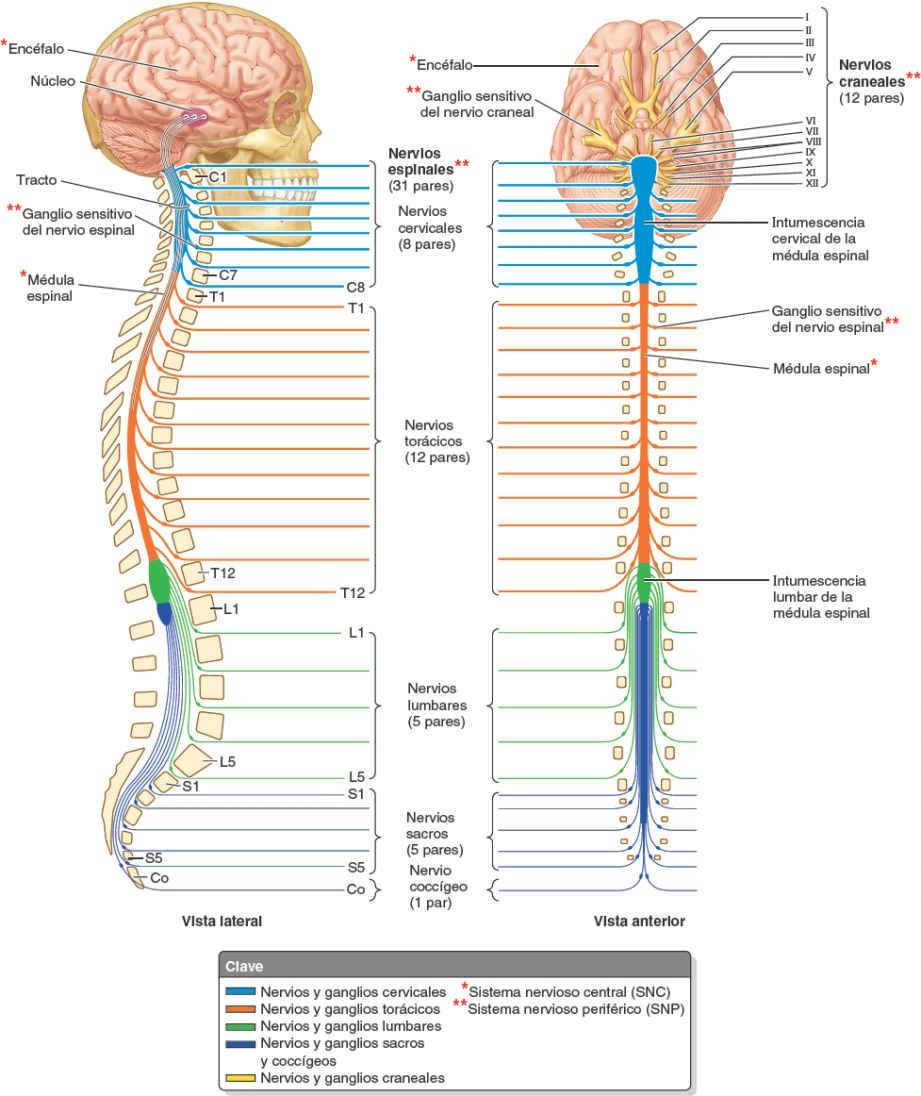
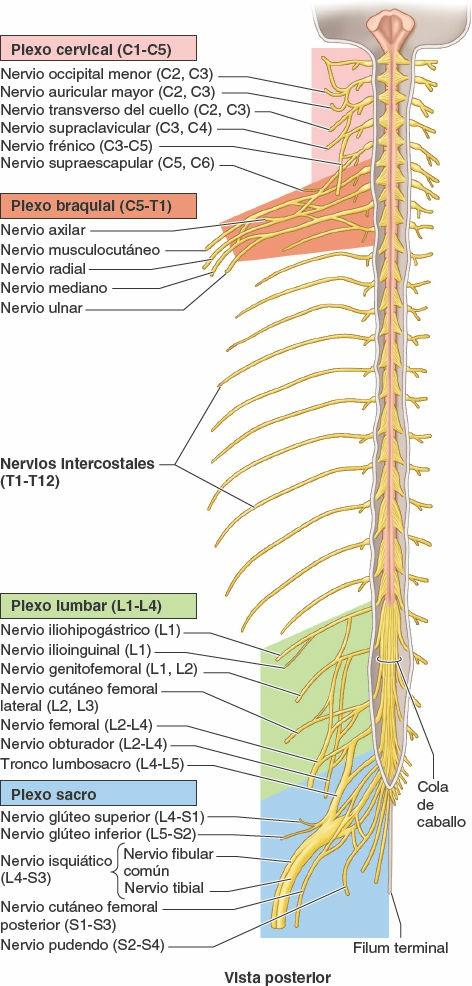
Los nervios se dividen en craneales o espinales, o derivados de ellos (fig. 1-30):
- Los nervios craneales salen de la cavidad craneal a través de los forámenes del cráneo, y se identifican por su nombre descriptivo (ej. «nervio troclear») o por un número romano (ej. «NC IV»). Sólo 11 de los 12 pares de nervios craneales se originan en el encéfalo; el par restante (NC XI) surge de la parte superior de la médula espinal.
- Los nervios espinales (segmentarios) salen de la columna vertebral a través de los forámenes intervertebrales. Los nervios espinales surgen en pares bilaterales desde un segmento específico de la médula espinal. Los 31 segmentos medulares y los 31 pares de nervios que surgen de ellos se identifican por una letra y un número (ej. «T4») para designar la región de la médula espinal y su orden de superior a inferior (C, cervical; T, torácica; L, lumbar; S, sacra, y Co, coccígea).
Nervios espinales
Los nervios espinales se inician en la médula espinal en forma de raicillas o filetes radiculares (un detalle que suele omitirse en los diagramas, en aras de la simplicidad), que convergen para formar dos raíces (fig. 1-34):
- Una raíz nerviosa anterior (ventral) compuesta por fibras motoras (eferentes) que viajan desde los cuerpos de las neuronas del cuerno (asta) anterior de la sustancia gris medular hasta los órganos efectores periféricos.
- Una raíz nerviosa posterior (dorsal) formada por fibras sensitivas (aferentes) desde los cuerpos celulares en los ganglios sensitivos de los nervios espinales o de la raíz posterior (dorsal) que se extienden periféricamente a las terminaciones sensitivas y centralmente al cuerno (asta) posterior de la sustancia gris medular.
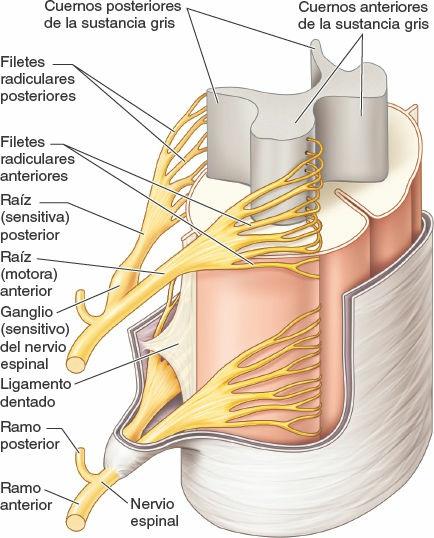
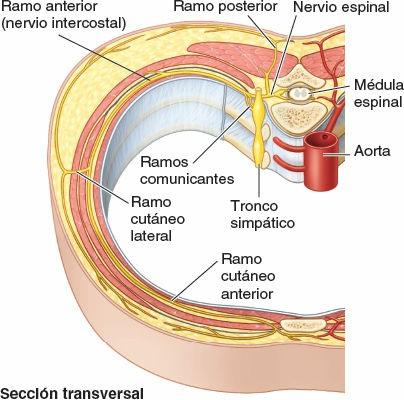
Las raíces nerviosas anterior y posterior se unen, dentro del foramen intervertebral o en sus inmediaciones cercanas, para formar un nervio espinal mixto (motor y sensitivo) que se divide enseguida en dos ramos: un ramo posterior (dorsal) y un ramo anterior (ventral). Al ser ramos del nervio espinal mixto, los ramos posterior y anterior llevan fibras motoras y sensitivas, al igual que todas sus ramificaciones subsiguientes. Los términos nervio motor y nervio sensitivo son casi siempre relativos y hacen referencia a la mayoría de los tipos de fibras que componen ese nervio. Los nervios que inervan los músculos del tronco o de los miembros (nervios motores) contienen también un 40% de fibras sensitivas, que conducen la sensibilidad dolorosa y la información propioceptiva. A la inversa, los nervios cutáneos (sensitivos) contienen fibras motoras que inervan las glándulas sudoríparas y los músculos lisos de los vasos sanguíneos y de los folículos pilosos.
La relación entre los nervios, la piel y el músculo se establece durante el desarrollo inicial. Ciertamente, la estructura segmentaria y la organización de los humanos no es tan evidente como en los anélidos, pero puede constatarse durante un período del desarrollo conocido como período somítico. Después de este período embrionario temprano, nuestra estructura segmentaria es más evidente en el esqueleto (vértebras y costillas), los nervios y los músculos de la región torácica.
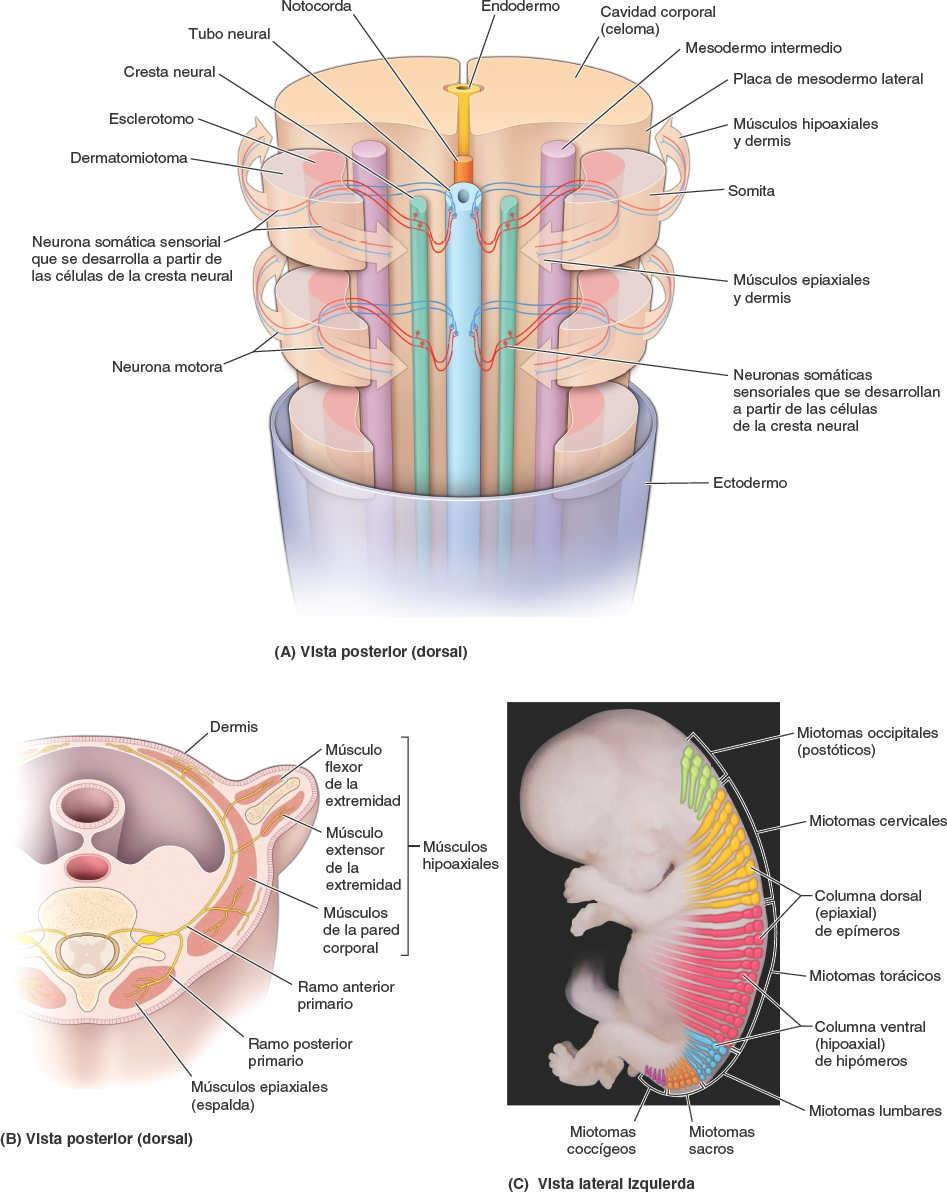
Durante el período somítico (fig. 1-35), el tejido que dará origen a los músculos, huesos y otros tejidos conectivos (incluyendo la dermis de la piel), tiene la apariencia de una fila bilateral de formaciones parecidas a galletas que rodean la médula espinal primitiva (tubo neural). Estas formaciones se denominan somitas:
- Las caras mediales de las somitas se convierten en esclerotomos, células que salen de las somitas y migran medialmente (fig. 1-35 A).
- Las células del esclerotomo que migran anteriormente rodean a la notocorda, formando los primordios de los cuerpos vertebrales.
- Las células del esclerotomo que migran dorsalmente rodean al tubo neural y forman los primordios del arco neural de las vértebras.
- La vista lateral de las somitas (dermatomiotomas) da origen a los músculos esqueléticos y la dermis de la piel.
- Las células del dermatomiotoma que migran en sentido posterior dan origen a los músculos intrínsecos o epiaxiales (profundos) de la espalda y la dermis que los cubre (fig. 1-35 B y C).
- Las células que migran en sentido anterior dan origen a los músculos hipoaxiales del tronco anterolateral y las extremidades, así como la dermis asociada.
- Los nervios se desarrollan en pares bilaterales que inervan al tejido que forma la dermis y el músculo de las somitas adyacentes (fig. 1-35 A).
- Las neuronas motoras que se desarrollan en el tubo neural anterior envían procesos periféricos hacia las regiones posterior y anterior del dermatomiotoma.
- Las neuronas sensoriales que se desarrollan en las crestas neurales envían procesos periféricos hacia estas regiones del dermatomiotoma y procesos centrales hacia la parte posterior del tubo neural.
- Las fibras nerviosas somáticas sensoriales y motoras que se organizan segmentariamente a lo largo del tubo neural forman parte de todos los nervios espinales y algunos nervios craneales.
- Los acúmulos de cuerpos celulares sensoriales derivados de la cresta neural, localizados fuera del SNC, forman los ganglios sensoriales.
- La relación entre los nervios y el tejido derivado del dermatomiotoma continúa durante toda la vida:
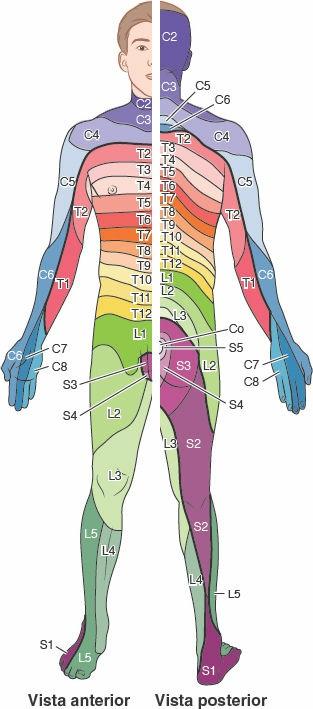
- El área unilateral de la piel inervada por un solo (parte derecha o izquierda de un par de) nervio espinal se llama dermatoma.
- La masa muscular unilateral inervada por un solo nervio espinal se denomina miotoma.
Durante toda la vida, la sección de un nervio espinal denervará el área de la piel y la masa muscular que nutría originalmente.
A partir de estudios clínicos de lesiones de las raíces posteriores o de los nervios espinales se han desarrollado mapas de los dermatomas para indicar el patrón típico de inervación de la piel por cada nervio espinal (fig. 1-36). Sin embargo, la lesión de una raíz posterior o de un nervio espinal raras veces daría lugar a entumecimiento en el área de la piel demarcada para ese nervio en dichos mapas, pues las fibras que transportan los nervios espinales adyacentes se solapan casi por completo al distribuirse por la piel, lo que proporciona una especie de doble cobertura. Por lo tanto, sería preferible que las líneas que delimitan los dermatomas en los mapas se difuminaran o se emplearan gradaciones de color. En general, han de interrumpirse al menos dos nervios espinales (o raíces posteriores; fig. 1-44 B) adyacentes para producir un área discernible de entumecimiento.
Al salir de los forámenes intervertebrales, los nervios espinales se dividen en dos ramos (figs. 1-37 y 1-44 B):
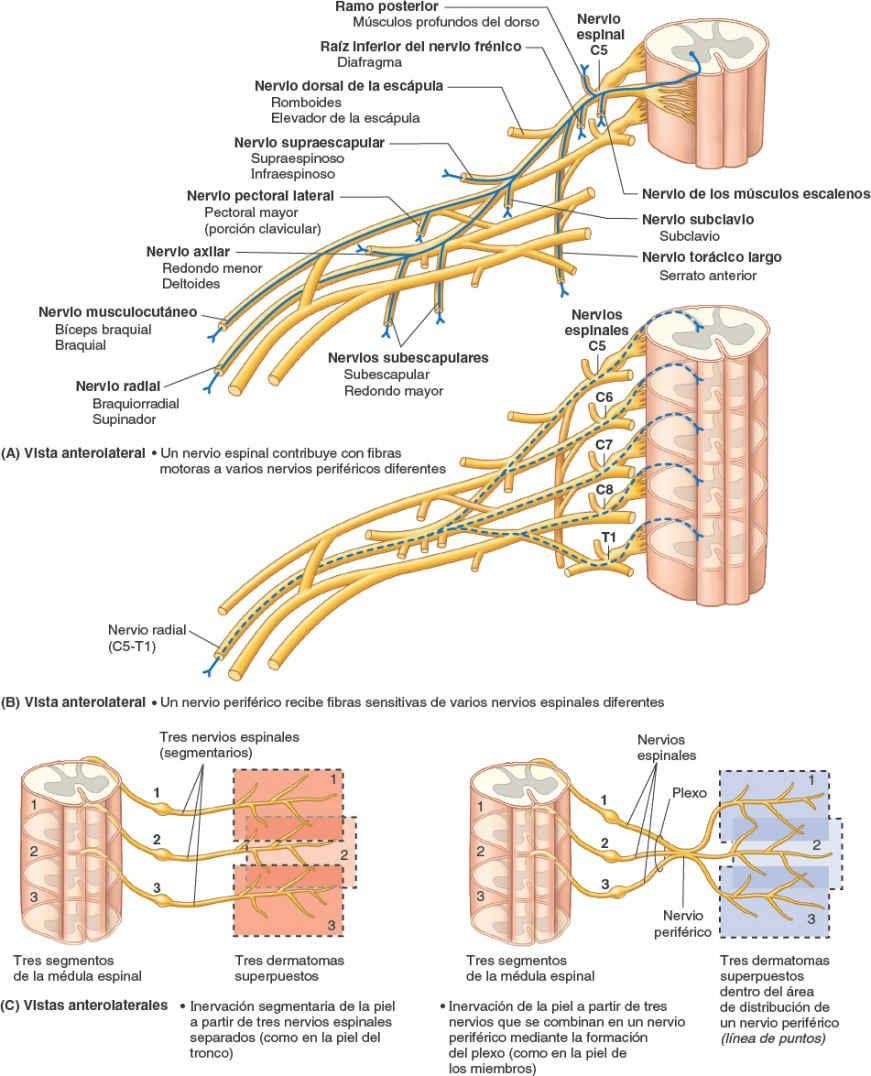
- Los ramos posteriores (primarios) de los nervios espinales aportan fibras nerviosas a las articulaciones sinoviales de la columna vertebral, a los músculos profundos del dorso y a la piel suprayacente, en un patrón segmentario. Como regla general, los ramos posteriores permanecen separados entre sí (no se unen para formar plexos nerviosos somáticos importantes).
- Los ramos anteriores (primarios) de los nervios espinales aportan fibras nerviosas al área restante, mucho más amplia, que consisten en la piel y los músculos hipoaxiales de las regiones anterior y lateral formada por las regiones anterior y lateral del tronco y los miembros superiores e inferiores. Los ramos anteriores que se distribuyen exclusivamente por el tronco permanecen en general separados entre sí e inervan también los músculos y la piel en un patrón segmentario (figs. 1-38, 1-39 y 1-44 B). En cambio, sobre todo en cuanto a la inervación de los miembros, la mayoría de los ramos anteriores emergen con uno o más ramos anteriores adyacentes y forman los plexos (redes) nerviosos somáticos principales, en los cuales se entremezclan sus fibras y de los que emerge una nueva serie de nervios periféricos multisegmentarios (figs. 1-39 y 1-40 A y B). Los ramos anteriores de los nervios espinales que participan en la formación de los plexos aportan fibras a múltiples nervios periféricos que se originan en el plexo (fig. 1-40 A); de manera recíproca, la mayoría de los nervios periféricos que surgen de los plexos contienen fibras de múltiples nervios espinales (fig. 1-40 B).
Aunque los nervios espinales pierden su identidad al dividirse y mezclarse en el plexo, las fibras que surgen de un determinado segmento medular y transcurren por un solo nervio espinal se distribuyen finalmente por un dermatoma segmentario, aunque pueden alcanzarlo mediante un nervio periférico multisegmentario que surge del plexo, que también lleva fibras a todos los dermatomas adyacentes o parte de ellos (fig. 1-40 C).
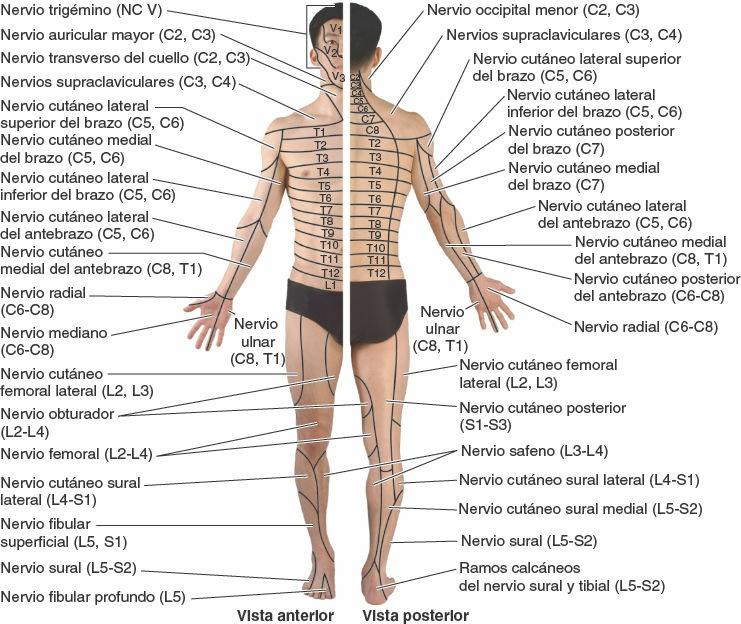
Por lo tanto, es importante distinguir entre la distribución de las fibras transportadas por los nervios espinales (inervación o distribución segmentaria, es decir, los dermatomas y miotomas etiquetados con una letra y un número, como «T4») y la de las fibras transportadas por los ramos de un plexo (inervación o distribución de los nervios periféricos, que se etiquetan con los nombres de los nervios periféricos, como el «nervio mediano») (figs. 1-36 y 1-38). Los mapas correspondientes a la inervación segmentaria (dermatomas, determinados por la experiencia clínica) o a la distribución de los nervios periféricos (determinada al disecar distalmente los ramos de un nervio) son completamente diferentes excepto en la mayor parte del tronco, donde al no haber plexos las distribuciones segmentaria y periférica son iguales. El solapamiento en la distribución cutánea de las fibras nerviosas conducidas por los nervios espinales adyacentes también tiene lugar en la distribución cutánea de las fibras nerviosas conducidas por los nervios periféricos adyacentes.
Nervios craneales
Al surgir del SNC, algunos nervios craneales llevan solamente fibras sensitivas, otros sólo llevan fibras motoras y otros son portadores de una mezcla de ambos tipos de fibras (fig. 1-41). Puesto que existen comunicaciones entre los nervios craneales, y entre estos y los nervios cervicales (espinales) superiores, un nervio que inicialmente sólo lleva fibras motoras puede recibir fibras sensitivas durante su curso distal, y viceversa. Excepto en los dos primeros nervios (que intervienen en los sentidos del olfato y la vista), los nervios craneales que llevan fibras sensitivas al encéfalo poseen ganglios sensitivos (similares a los ganglios sensitivos de los nervios espinales o de las raíces posteriores), donde se localizan los cuerpos celulares de las fibras seudomonopolares. Aunque, por definición, el término dermatoma se aplica solamente a los nervios espinales, pueden identificarse áreas similares de piel inervada por un determinado nervio craneal y elaborar mapas de esta distribución. Sin embargo, a diferencia de los dermatomas, hay poco solapamiento en la inervación de las zonas cutáneas inervadas por los nervios craneales.
Fibras somáticas y viscerales
Los tipos de fibras transportadas por los nervios craneales o espinales son los siguientes (fig. 1-41):
- Fibras somáticas:
- Fibras sensitivas generales (fibras aferentes somáticas generales), que transmiten las sensaciones corporales al SNC; pueden ser sensaciones exteroceptivas de la piel (dolor, temperatura, tacto y presión) o dolorosas, y sensaciones propioceptivas de los músculos, tendones y articulaciones. Las sensaciones propioceptivas suelen ser subconscientes y proporcionan información sobre la posición de las articulaciones y la tensión de los tendones y músculos. Esta información se combina con la procedente del aparato vestibular del oído interno para conocer la orientación espacial del cuerpo y los miembros, independientemente de la información visual.
- Fibras motoras somáticas (fibras eferentes somáticas generales), que transmiten impulsos a los músculos esqueléticos (voluntarios).
- Fibras viscerales:
- Fibras sensitivas viscerales (fibras aferentes viscerales generales), que transmiten las sensaciones reflejas viscerales dolorosas o subconscientes (ej. información sobre distensión, gases en sangre y presión arterial) de los órganos huecos y los vasos sanguíneos, que llegan al SNC.
- Fibras motoras viscerales (fibras eferentes viscerales generales), que transmiten impulsos a los músculos lisos (involuntarios) y a los tejidos glandulares. Dos tipos de fibras, presinápticas y postsinápticas, actúan conjuntamente para conducir los impulsos del SNC a los músculos lisos o a las glándulas.
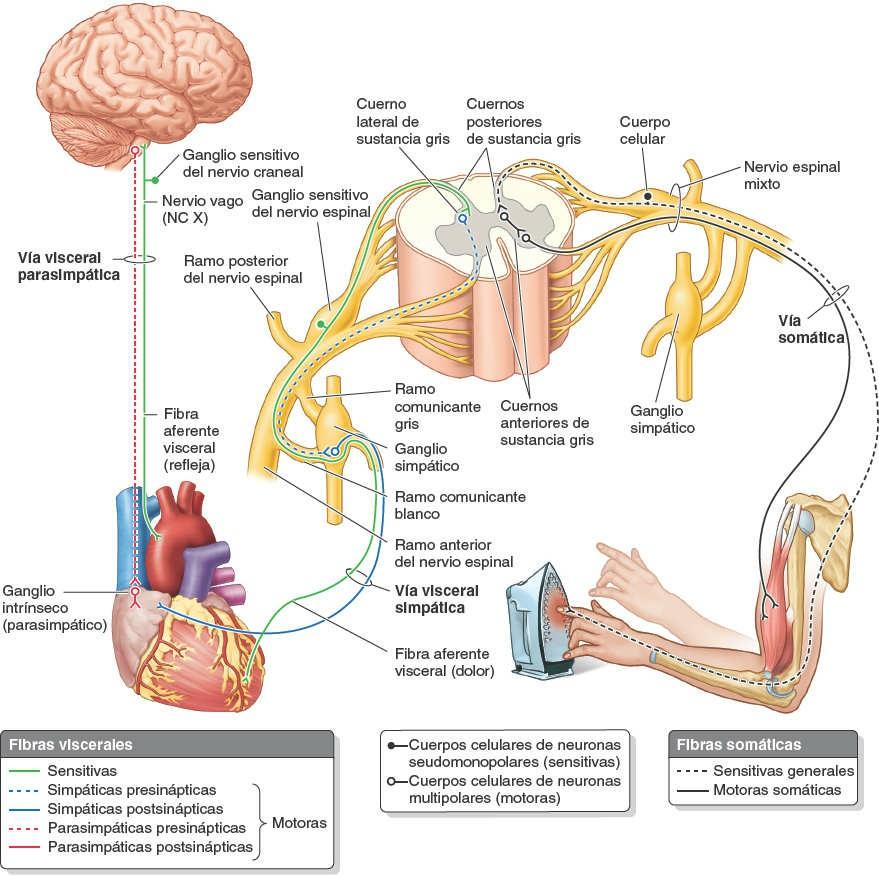
Ambos tipos de fibras sensitivas (viscerales y generales) son prolongaciones de las neuronas seudomonopolares, cuyos cuerpos celulares se hallan fuera del SNC, en los ganglios sensitivos de los nervios espinales o craneales (figs. 1-41 y 1-42). Las fibras motoras de los nervios son axones de las neuronas multipolares. Los cuerpos de las neuronas motoras somáticas y presinápticas viscerales se localizan en la sustancia gris de la médula espinal. Los cuerpos de las neuronas motoras postsinápticas se hallan fuera del SNC, en los ganglios autónomos.
Además de las clases de fibras citadas, algunos nervios craneales llevan también fibras sensitivas especiales para determinados sentidos (olfato, vista, oído, equilibrio y gusto). Basándose en el origen embrionario/filogenético de ciertos músculos de la cabeza y del cuello, algunas fibras motoras conducidas por los nervios craneales a los músculos estriados se han calificado tradicionalmente como «viscerales especiales»; pero, como la designación es confusa y no se aplica en clínica, aquí se omitirá. Estas fibras se designan a veces como motoras branquiales, como referencia al tejido muscular derivado de los arcos faríngeos embrionarios.
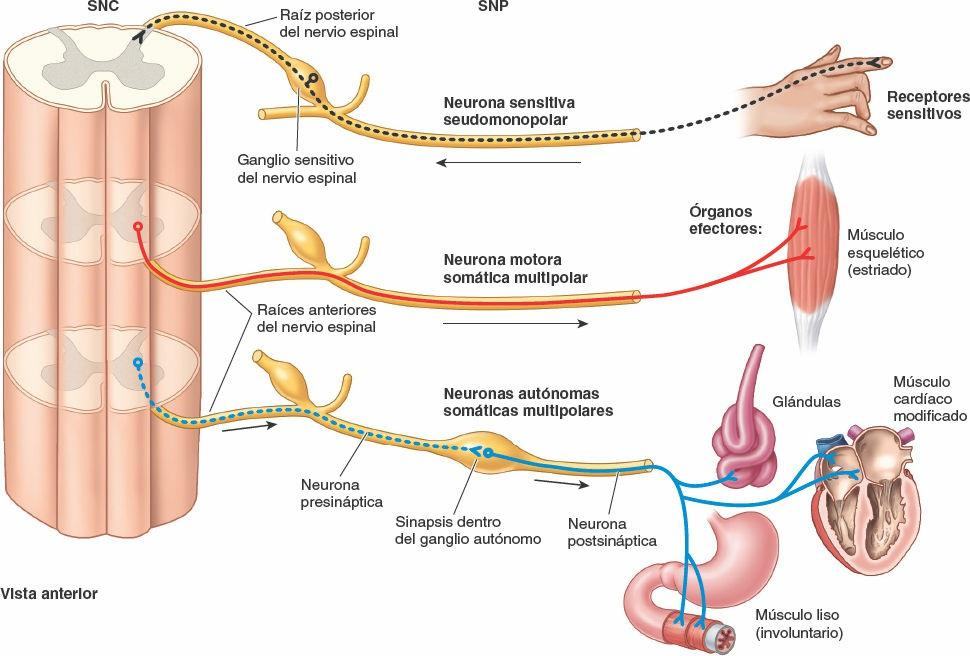
Sistema nervioso somático
El sistema nervioso somático está compuesto por las porciones somáticas del SNC y el SNP. Proporciona inervación sensitiva y motora a todas las partes del cuerpo (del griego soma), excepto a las vísceras de las cavidades corporales, el músculo liso y las glándulas (figs. 1-41 y 1-42). El sistema somático sensitivo transmite las sensaciones de dolor, temperatura y posición desde los receptores sensitivos. La mayoría de estas sensaciones alcanzan niveles conscientes (las notamos). El sistema somático motor inerva sólo los músculos esqueléticos, con estimulación de los movimientos voluntarios y reflejos, mediante una contracción muscular como ocurre en respuesta, por ejemplo, al tocar una plancha caliente.
Sistema nervioso autónomo
El sistema nervioso autónomo (SNA), denominado clásicamente sistema nervioso visceral o sistema motor visceral (figs. 1-41 y 1-42), se compone de fibras motoras que estimulan el músculo liso (involuntario), el músculo cardíaco modificado (estimulación intrínseca y tejido de conducción cardíaco) y las células glandulares (secretoras). Sin embargo, las fibras eferentes viscerales del SNA van acompañadas de fibras aferentes viscerales. Como componente aferente de los reflejos autónomos y por conducir los impulsos del dolor visceral, estas fibras aferentes también tienen un papel regulador de la función visceral.
Las fibras nerviosas eferentes y los ganglios del SNA están organizados en dos sistemas o divisiones: la división simpática (toracolumbar) y la división parasimpática (craneosacra). A diferencia de las inervaciones sensitiva y motora somática, en las cuales interviene una sola neurona en el paso de los impulsos entre el SNC y las terminaciones sensitivas o el órgano efector, en ambas divisiones del SNA interviene una serie de dos neuronas multipolares para conducir los impulsos desde el SNC al órgano efector (fig. 1-42). El cuerpo celular de la primera neurona presináptica (preganglionar) está localizado en la sustancia gris del SNC. Su fibra (axón) establece sinapsis sólo en el cuerpo celular de una neurona postsináptica (posganglionar), la segunda neurona de la serie. Los cuerpos celulares de estas segundas neuronas se hallan fuera del SNC, en los ganglios autónomos, con fibras que finalizan en el órgano efector (músculo liso, músculo cardíaco modificado o glándulas).
La distinción anatómica entre las divisiones simpática y parasimpática del SNA se basa principalmente en:
- La localización de los cuerpos celulares presinápticos.
- La identidad de los nervios que conducen las fibras presinápticas desde el SNC.
Una distinción funcional de importancia farmacológica para la práctica médica consiste en que las neuronas postsinápticas de las dos divisiones liberan generalmente diferentes sustancias neurotransmisoras: noradrenalina en la división simpática (excepto en las glándulas sudoríparas) y acetilcolina en la división parasimpática.
División simpática (toracolumbar) del SNA
Los cuerpos celulares de las neuronas presinápticas de la división simpática del SNA se hallan en un solo lugar: las columnas celulares o núcleos intermediolaterales (IML) de la médula espinal (fig. 1-43). Los núcleos IML pares (derecho e izquierdo) forman parte de la sustancia gris de los segmentos torácicos (T1-T12) y lumbares altos (L1-L2 o L3) de la médula espinal (de aquí la denominación alternativa «toracolumbar» para esta división). En los cortes transversales de esta parte de la médula espinal, los núcleos IML aparecen como pequeños cuernos laterales en la H que forma la sustancia gris, como una prolongación de la barra transversal de la H entre los cuernos posterior y anterior. Los núcleos IML se hallan organizados somatotópicamente (es decir, dispuestos de tal modo que los cuerpos celulares que intervienen en la inervación de la cabeza están situados en la parte superior, y los que se ocupan de inervar las vísceras pelvianas y los miembros inferiores se localizan en la parte inferior). De este modo, es posible deducir la localización de los cuerpos celulares simpáticos presinápticos que inervan una determinada parte del cuerpo.
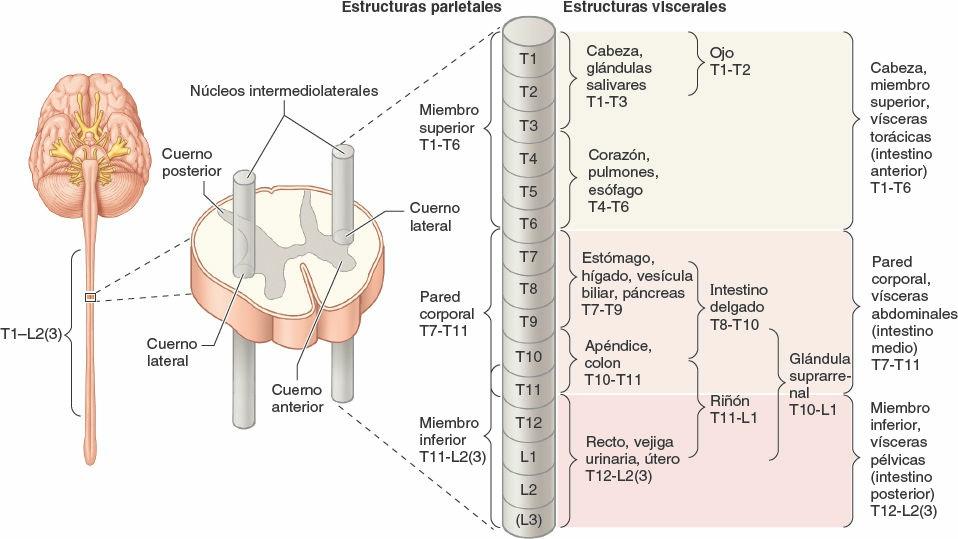
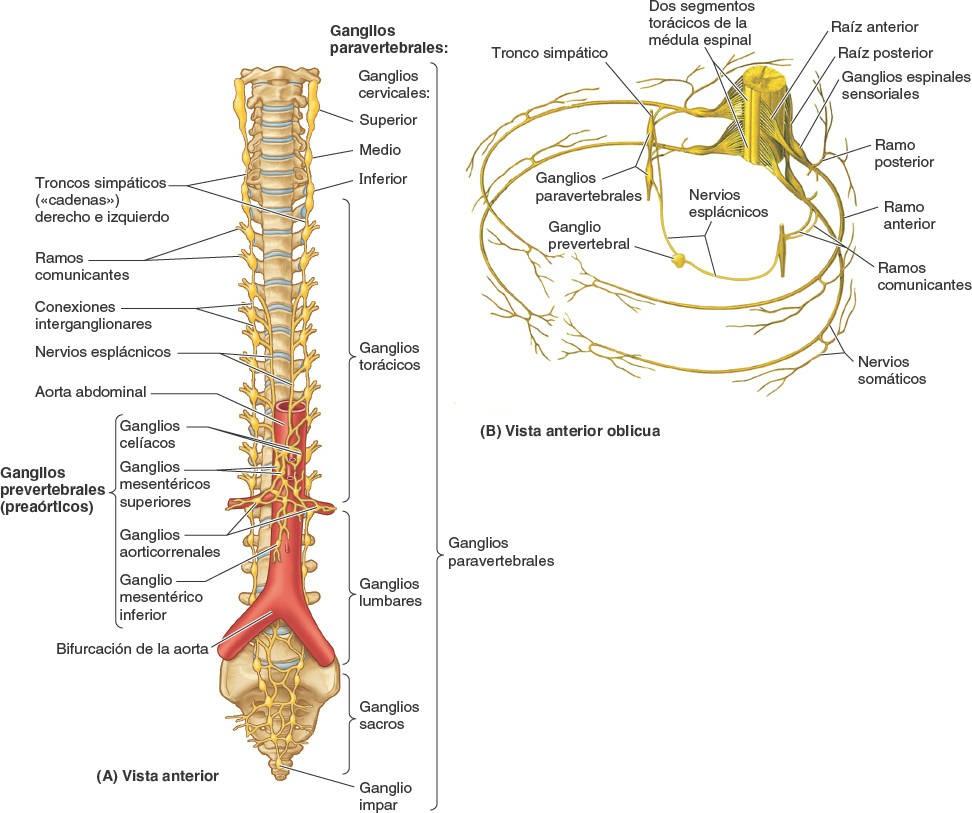
Los cuerpos celulares de las neuronas postsinápticas del sistema nervioso simpático se encuentran en dos localizaciones, los ganglios paravertebrales y prevertebrales (fig. 1-44):
- Los ganglios paravertebrales están unidos para formar los troncos (cadenas) simpáticos derecho e izquierdo a cada lado de la columna vertebral y se extienden a lo largo de esta. El ganglio paravertebral superior (el ganglio cervical superior de cada tronco simpático) está situado en la base del cráneo. El ganglio impar se forma en la parte inferior, donde se unen los dos troncos a nivel del cóccix.
- Los ganglios prevertebrales se hallan en los plexos que rodean los orígenes de las ramas principales de la aorta abdominal (de las que toman su nombre), como los dos grandes ganglios celíacos que rodean el origen del tronco celíaco (una arteria principal que nace de la aorta).
Debido a que son fibras motoras, los axones de las neuronas presinápticas abandonan la médula espinal a través de las raíces anteriores y penetran en los ramos anteriores de los nervios espinales T1-L2 o L3 (figs. 1-45 y 1-46). Casi inmediatamente después de penetrar, todas las fibras simpáticas presinápticas abandonan los ramos anteriores de estos nervios espinales y pasan a los troncos simpáticos a través de los ramos comunicantes blancos. Dentro de los troncos simpáticos, las fibras presinápticas siguen uno de cuatro recorridos posibles:
- Ascienden en el tronco simpático para establecer sinapsis con una neurona postsináptica de un ganglio paravertebral más superior.
- Descienden en el tronco simpático para establecer sinapsis con una neurona postsináptica de un ganglio paravertebral más inferior.
- Entran y establecen sinapsis inmediatamente con una neurona postsináptica del ganglio paravertebral situado a ese nivel.
- Pasan a través del tronco simpático sin establecer sinapsis y siguen por un nervio esplácnico abdominopélvico (un ramo del tronco que interviene en la inervación de las vísceras abdominopélvicas) para alcanzar los ganglios prevertebrales.
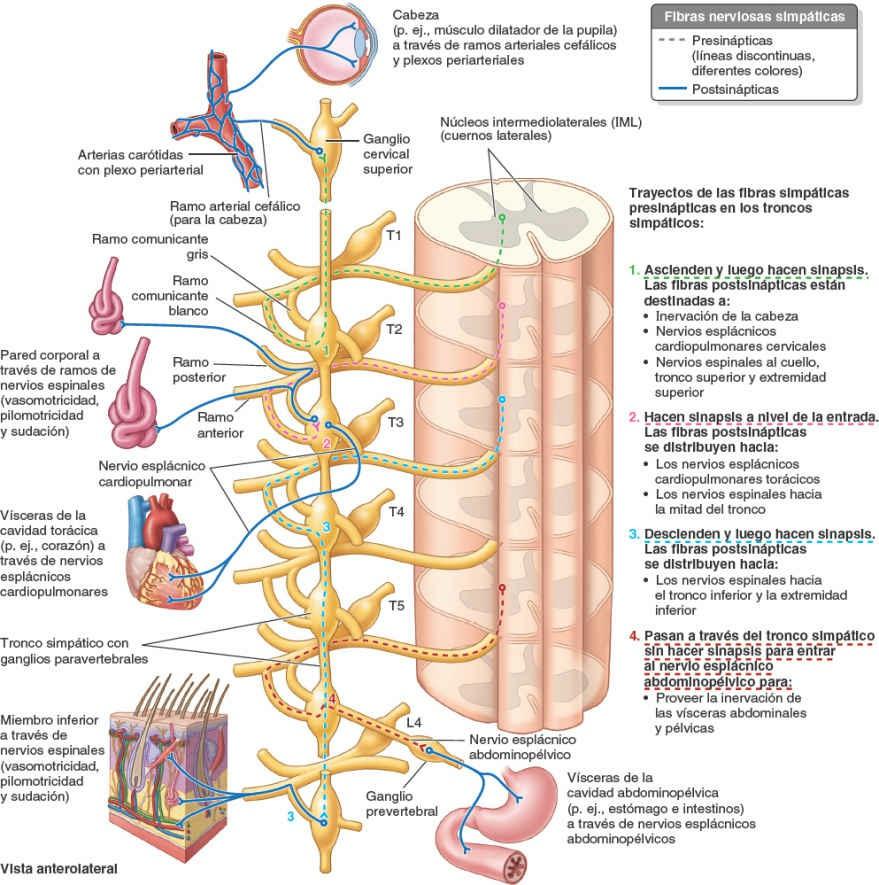
Las fibras simpáticas presinápticas que proporcionan inervación autónoma a la cabeza, el cuello, la pared corporal, los miembros y la cavidad torácica, siguen una de las tres primeras vías y establecen sinapsis con los ganglios paravertebrales. Las fibras simpáticas presinápticas que inervan las vísceras de la cavidad abdominopélvica siguen la cuarta vía.
Las fibras simpáticas postsinápticas superan en gran número a las presinápticas: cada fibra simpática presináptica establece sinapsis con 30 o más fibras postsinápticas. Las fibras simpáticas postsinápticas que se distribuyen por el cuello, la pared corporal y los miembros, pasan desde los ganglios paravertebrales de los troncos simpáticos a los ramos anteriores adyacentes de los nervios espinales, a través de los ramos comunicantes grises (fig. 1-46). Por este medio penetran en todos los ramos de los 31 pares de nervios espinales, incluidos los ramos posteriores.
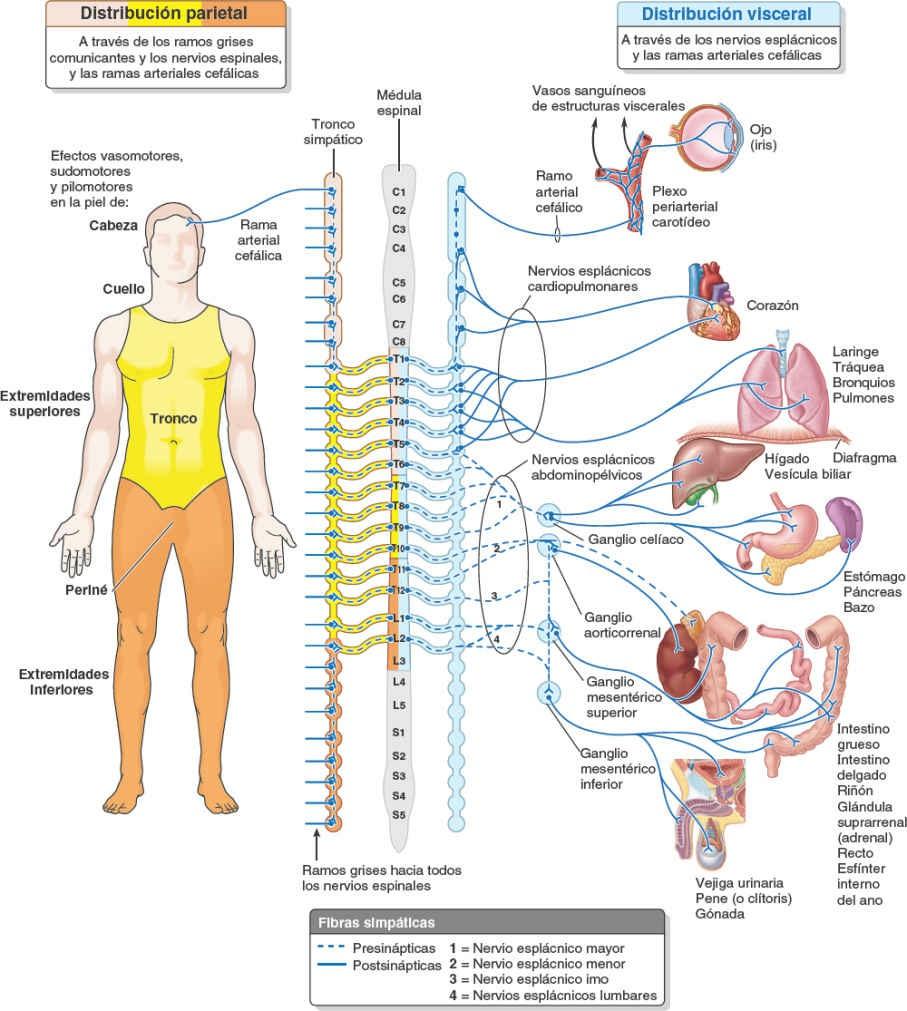
Las fibras simpáticas postsinápticas estimulan la contracción de los vasos sanguíneos (vasomotricidad) y los músculos erectores del pelo (pilomotricidad, «piel de gallina»); además, provocan la sudación. Todas las fibras simpáticas postsinápticas que realizan estas funciones en la cabeza (más la inervación del músculo dilatador del iris) tienen sus cuerpos celulares en el ganglio cervical superior, situado en el extremo superior del tronco simpático. Desde dicho ganglio, pasan a través de una rama arterial cefálica para formar plexos nerviosos periarteriales que siguen las ramas de las arterias carótidas, o pasan directamente a los nervios craneales próximos, para alcanzar su punto de destino en la cabeza.
Los nervios esplácnicos llevan fibras eferentes (autónomas) y aferentes viscerales hacia y desde las vísceras de las cavidades corporales. Las fibras simpáticas postsinápticas destinadas a las vísceras de la cavidad torácica (ej. corazón, pulmones y esófago) pasan a través de los nervios esplácnicos cardiopulmonares y penetran en los plexos cardíaco, pulmonar y esofágico (figs. 1-45 y 1-46). Las fibras simpáticas presinápticas que inervan las vísceras de la cavidad abdominopélvica (ej. estómago e intestinos) pasan a los ganglios prevertebrales a través de los nervios esplácnicos abdominopélvicos (que componen los nervios esplácnicos torácicos mayor, menor, imo y esplácnicos lumbares) (figs. 1-45 a 1-47).
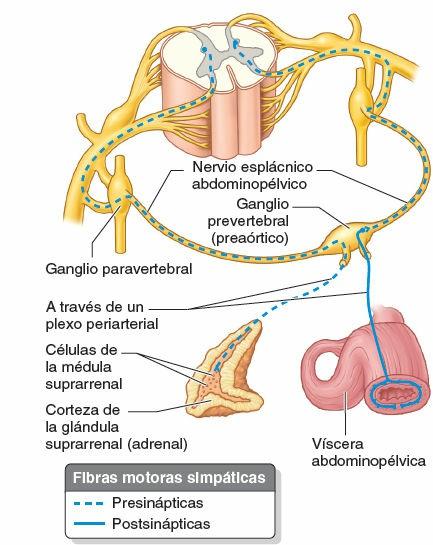
Todas las fibras simpáticas presinápticas de los nervios esplácnicos abdominopélvicos, excepto los que inervan las glándulas suprarrenales, establecen sinapsis en ganglios prevertebrales. Las fibras postsinápticas de los ganglios prevertebrales forman plexos periarteriales que siguen las ramas de la aorta abdominal para alcanzar sus puntos de destino.
Algunas fibras simpáticas presinápticas pasan a través de los ganglios prevertebrales celíacos sin establecer sinapsis y siguen para terminar directamente en las células de la médula suprarrenal (fig. 1-47). Estas células actúan como un tipo especial de neurona postsináptica que, en vez de liberar su sustancia neurotransmisora sobre las células de un determinado órgano efector, la liberan al torrente sanguíneo para que circule por todo el cuerpo y produzca una respuesta simpática generalizada. Por lo tanto, la inervación simpática de esta glándula es excepcional.
Como se ha dicho, las fibras simpáticas postsinápticas son componentes de prácticamente todos los ramos de los nervios espinales. Por este medio, y por vía de los plexos periarteriales, se extienden e inervan todos los vasos sanguíneos del organismo (la función primaria del sistema nervioso simpático), así como las glándulas sudoríparas, los músculos erectores del pelo y las estructuras viscerales. De este modo, el sistema nervioso simpático llega prácticamente a todas las partes del cuerpo, con la rara excepción de ciertos tejidos avasculares como el cartílago y las uñas. Debido a que las dos series de ganglios simpáticos (paravertebrales y prevertebrales) están situadas centralmente en el organismo y próximas a la línea media (y por lo tanto bastante cercanas a la médula espinal), las fibras presinápticas de esta división son relativamente cortas, mientras que las fibras postsinápticas son relativamente largas, al llegar a todas las partes del cuerpo.
División parasimpática (craneosacra) del SNA
Los cuerpos de las neuronas parasimpáticas presinápticas están situados en dos partes del SNC, y sus fibras salen por dos vías. Esta disposición es la causa de la denominación alternativa «craneosacra» para referirse a la división parasimpática del SNA (fig. 1-48):
- En la sustancia gris del tronco del encéfalo, las fibras salen del SNC dentro de los nervios craneales III, VII, IX y X; estas fibras constituyen la eferencia parasimpática craneal.
- En la sustancia gris de los segmentos sacros de la médula espinal (S2-S4), las fibras salen del SNC a través de las raíces anteriores de los nervios espinales sacros S2-S4 y los nervios esplácnicos pélvicos que se originan de sus ramos anteriores; estas fibras constituyen la eferencia parasimpática sacra.
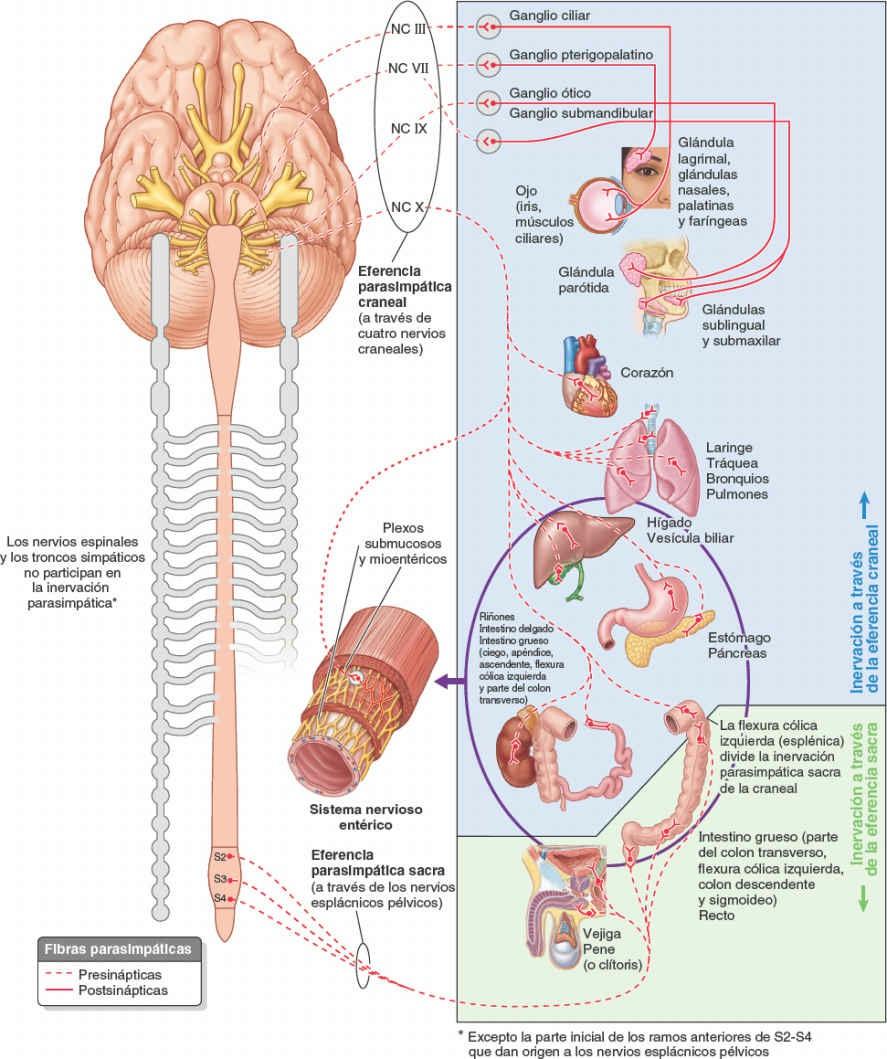
Como es de esperar, la eferencia craneal proporciona inervación parasimpática a la cabeza, y la eferencia sacra a las vísceras pélvicas. Sin embargo, en términos de inervación de las vísceras torácicas y abdominales domina la eferencia craneal a través del nervio vago (NC X). Este proporciona inervación para todas las vísceras torácicas y la mayoría del tracto gastrointestinal desde el esófago hasta la mayor parte del intestino grueso (hasta su flexura cólica izquierda).
La eferencia sacra para el tracto gastrointestinal inerva sólo el colon descendente y sigmoideo y el recto.
Sin tener en cuenta la extensa influencia de su eferencia craneal, el sistema parasimpático está mucho más restringido en su distribución que el sistema simpático. El sistema parasimpático sólo se distribuye hacia la cabeza, las cavidades viscerales del tronco y los tejidos eréctiles de los genitales externos. Con excepción de estos últimos, no alcanza las paredes corporales ni los miembros, y excepto las partes iniciales de los ramos anteriores de los nervios espinales S2-S4, sus fibras no forman parte de los nervios espinales ni de sus ramos.
En la cabeza se encuentran cuatro pares de ganglios parasimpáticos. En otros lugares, las fibras parasimpáticas presinápticas hacen sinapsis con cuerpos celulares postsinápticos, que se encuentran aislados en la pared del órgano inervado o sobre ella (ganglios intrínsecos o entéricos). En consecuencia, en esta división, muchas fibras parasimpáticas presinápticas son largas y se extienden desde el SNC hasta el órgano efector, mientras que las fibras postsinápticas son cortas y discurren desde un ganglio localizado cerca del órgano efector o en él mismo.
Sistema nervioso entérico
Actualmente se sabe que las neuronas motoras que han sido identificadas como las neuronas parasimpáticas postsinápticas del tracto gastrointestinal desempeñan un papel mucho más sofisticado que simplemente recibir y transmitir la señal de las fibras parasimpáticas presinápticas hacia el músculo liso y las glándulas. Estas neuronas motoras son componentes importantes del sistema nervioso entérico (SNE) y cada vez más se han identificado como un tercer componente del sistema motor visceral o incluso un «segundo cerebro» debido a su complejidad, función integrativa y capacidad de funcionar de forma autónoma, sin conexión con el SNC a través de otras divisiones del SNA o las aferentes viscerales extrínsecas.
El SNE consta de dos plexos interconectados en las paredes del tracto gastrointestinal: el plexo mientérico de la pared muscular y el plexo submucoso, que se localiza por debajo del revestimiento intestinal o mucosa y la inerva (fig. 1-48, recuadro). Además de las neuronas motoras, que están extensamente interconectadas tanto directamente como entre las neuronas, el plexo incluye neuronas aferentes primarias intrínsecas que reciben un impulso local y estimulan a las neuronas motoras, formando circuitos reflejos locales que integran intrínsecamente la secreción exocrina y endocrina, el efecto vasomotor, la micromotilidad y la actividad inmunitaria del intestino. Esta actividad local sólo está modulada por el impulso proveniente de las fibras parasimpáticas extrínsecas y simpáticas. En el capítulo 5 se proporciona información más detallada sobre el SNE.
Funciones de las divisiones del SNA
Aunque los sistemas simpático y parasimpático inervan estructuras involuntarias (y a menudo influyen en ellas), sus efectos son diferentes, usualmente opuestos pero bien coordinados (figs. 1-46 y 1-48). En general, el sistema simpático es un sistema catabólico (con gasto energético) que permite al organismo afrontar el estrés, como al prepararse para la respuesta de lucha o fuga. El sistema parasimpático es principalmente un sistema homeostático o anabólico (con conservación de energía), que promueve los procesos tranquilos y ordenados del organismo, como los que permiten la alimentación y la asimilación. En la tabla 1-2 se resumen las funciones específicas del SNA y sus divisiones.
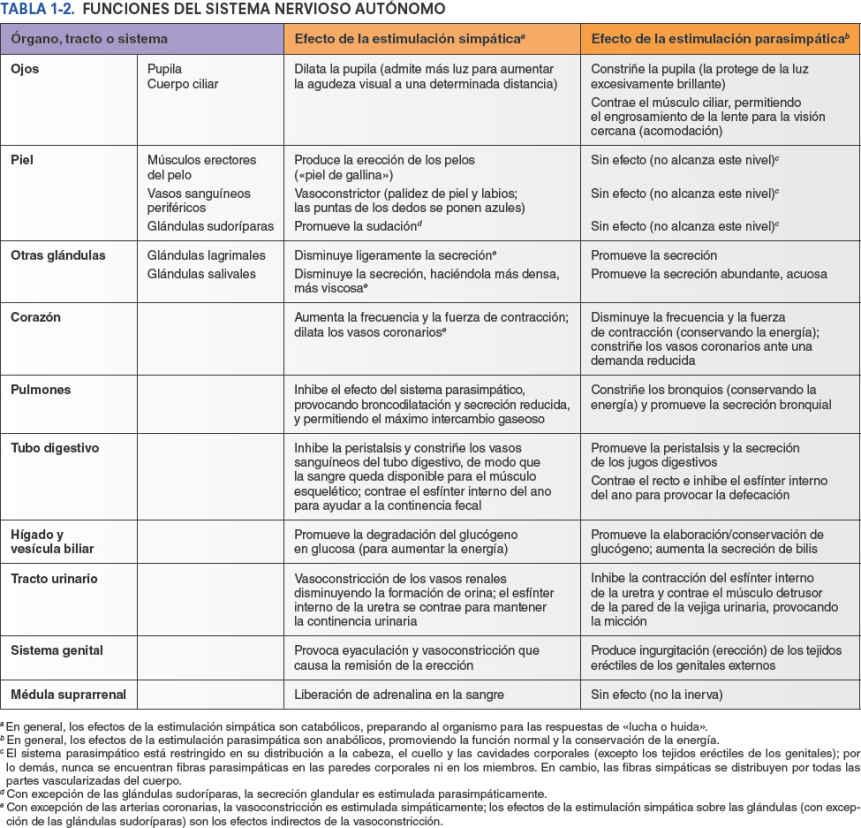
La función primaria del sistema simpático es regular los vasos sanguíneos, lo que se logra por varios medios, con diferentes consecuencias. Los vasos sanguíneos de todo el cuerpo reciben inervación tónica de los nervios simpáticos, con una moderada vasoconstricción basal. En la mayoría de los lechos vasculares, al aumentar las señales simpáticas se incrementa dicha vasoconstricción, y al descender se permite la vasodilatación. Sin embargo, en algunas partes del cuerpo las señales simpáticas son vasodilatadoras (es decir, las sustancias transmisoras simpáticas inhiben la vasoconstricción activa, lo cual permite que los vasos sanguíneos se dilaten pasivamente por la presión arterial). En los vasos coronarios, en los vasos de los músculos esqueléticos y en los genitales externos, la estimulación simpática produce vasodilatación.
Sensibilidad visceral
Las fibras aferentes viscerales poseen importantes relaciones en el SNA, tanto anatómicas como funcionales. Habitualmente no percibimos los impulsos sensitivos de estas fibras, que aportan información sobre el estado del medio interno del organismo. Esta información se integra en el SNC y a menudo desencadena reflejos viscerales o somáticos, o ambos. Los reflejos viscerales regulan la presión arterial y la bioquímica sanguínea, al modificar ciertas funciones como las frecuencias cardíaca y respiratoria y la resistencia vascular.
La sensibilidad visceral que alcanza el nivel de la consciencia se percibe generalmente en forma de dolor, mal localizado o como calambres, o con sensaciones de hambre, repleción o náusea. En las intervenciones practicadas con anestesia local, el cirujano puede manejar, seccionar, pinzar o incluso quemar (cauterizar) los órganos viscerales sin provocar sensaciones conscientes. En cambio, ciertas estimulaciones pueden provocar dolor:
- Distensión súbita.
- Espasmos o contracciones intensas.
- Irritantes químicos.
- Estimulación mecánica, sobre todo cuando el órgano se halla activo.
- Procesos patológicos (especialmente la isquemia) que disminuyen el umbral normal de estimulación.
La actividad normal no suele producir ninguna sensación, pero puede haberla cuando el aporte sanguíneo es insuficiente (isquemia). La mayoría de las sensaciones reflejas viscerales (inconscientes) y algunas sensaciones dolorosas viajan por las fibras aferentes viscerales que acompañan de forma retrógrada a las fibras parasimpáticas. Casi todos los impulsos de dolor visceral (desde el corazón y la mayoría de los órganos de la cavidad peritoneal) discurren centralmente a lo largo de las fibras aferentes viscerales que acompañan a las fibras simpáticas.