02. Pared Torácica
La pared torácica está formada por la caja torácica y los músculos que se extienden entre las costillas, así como por la piel, el tejido subcutáneo, los músculos y las fascias que cubren su cara anterolateral. Las mismas estructuras que cubren su cara posterior se consideran pertenecientes al dorso. Las glándulas mamarias de las mamas se localizan dentro del tejido subcutáneo de la pared torácica. Los músculos axioapendiculares anterolaterales que recubren la caja torácica y constituyen el lecho de la mama se encuentran en la pared torácica y pueden ser considerados parte de ella, pero son claramente músculos del miembro superior si nos atenemos a criterios funcionales y de inervación. Los mencionaremos brevemente aquí.
La forma abovedada de la caja torácica le proporciona una rigidez notable, considerando el escaso peso de sus componentes, lo que permite:
- Proteger los órganos vitales del tórax y del abdomen (la mayor parte contienen aire o líquido) frente a las fuerzas del exterior.
- Resistir las presiones negativas internas (subatmosféricas) que se generan por el retroceso elástico de los pulmones y por los movimientos de inspiración.
- Proporcionar inserción para los miembros superiores y sostener su peso.
- Proporcionar inserción (origen) a muchos de los músculos que mantienen la posición de los miembros superiores en relación con el tronco, así como a los músculos del abdomen, el cuello, el dorso y la respiración.
Aunque la forma de la caja torácica proporciona rigidez, sus articulaciones y la delgadez y la flexibilidad de las costillas le permiten absorber muchos golpes y compresiones externas sin fracturarse, y cambiar su forma para la respiración. Debido a que las estructuras internas más importantes del tórax (corazón, grandes vasos, pulmones y tráquea), así como su suelo y paredes, están en constante movimiento, el tórax es una de las regiones más dinámicas del cuerpo. Con cada respiración, los músculos de la pared torácica —trabajando conjuntamente con el diafragma y los músculos de la pared abdominal— varían el volumen de la cavidad torácica, primero expandiendo su capacidad, lo que produce la expansión de los pulmones y la entrada del aire, y a continuación, debido a la elasticidad pulmonar y la relajación muscular, disminuyendo el volumen de la cavidad y produciendo la salida del aire.
Esqueleto de la pared torácica
El esqueleto del tórax forma la caja torácica osteocartilaginosa (fig. 4-1), que protege las vísceras torácicas y algunos órganos abdominales. El esqueleto del tórax incluye 12 pares de costillas y los cartílagos costales asociados, 12 vértebras torácicas con los discos intervertebrales interpuestos entre ellas, y el esternón. Las costillas y los cartílagos costales forman la mayor parte de la caja torácica; ambos se identifican numéricamente, desde el más superior (1ª costilla o cartílago costal) hasta el más inferior (12ª).
Costillas, cartílagos costales y espacios intercostales
Las costillas son huesos planos y curvos que constituyen la mayor parte de la caja torácica (figs. 4-1 y 4-2). Son especialmente ligeras en cuanto a su peso, y muy elásticas. Cada costilla posee un interior esponjoso que contiene la médula ósea (tejido hematopoyético), productora de células sanguíneas. Hay tres tipos de costillas que pueden clasificarse como típicas o atípicas:
- Las costillas verdaderas (vertebrocostales, 1ª-7ª costillas) se unen directamente al esternón mediante sus propios cartílagos costales.
- Las costillas falsas (vertebrocondrales, las costillas 8ª, 9ª y normalmente la 10ª) tienen cartílagos que se unen al de la costilla inmediatamente superior a ella; de este modo, su conexión con el esternón es indirecta.
- Las costillas flotantes (libres, costillas 11ª, 12ª y a veces la 10ª) tienen cartílagos rudimentarios que nunca conectan, ni directa ni indirectamente, con el esternón; por el contrario, terminan en la musculatura posterior del abdomen.
Las costillas típicas (3ª-9ª) poseen los siguientes componentes:
- Una cabeza con forma de cuña y dos caras articulares separadas por la cresta de la cabeza de la costilla (figs. 4-2 y 4-3). Una cara se articula con su vértebra numéricamente correspondiente y la otra lo hace con la inmediatamente superior.
- Un cuello que conecta la cabeza con el cuerpo al nivel del tubérculo.
- Un tubérculo en la unión del cuello y el cuerpo. El tubérculo tiene una porción articular lisa para la articulación con el proceso (apófisis) transverso de la vértebra correspondiente, y una porción no articular rugosa para la inserción del ligamento costotransverso (fig. 4-8 B).
- Un cuerpo delgado, plano y curvo, especialmente en el ángulo de la costilla donde la costilla gira de forma anterolateral. El ángulo también constituye el límite lateral de la inserción en las costillas de los músculos profundos del dorso (figs. 2-29 y 2-31). En la superficie interna cóncava del cuerpo hay un surco de la costilla, paralelo al borde inferior de la costilla, que protege en parte al nervio y los vasos intercostales.
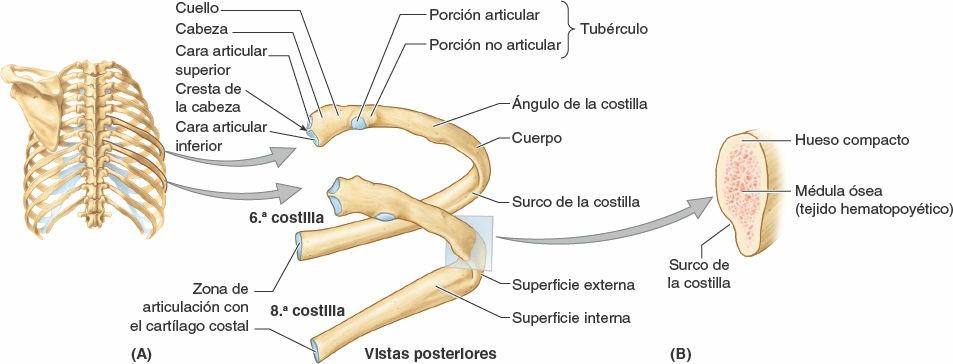
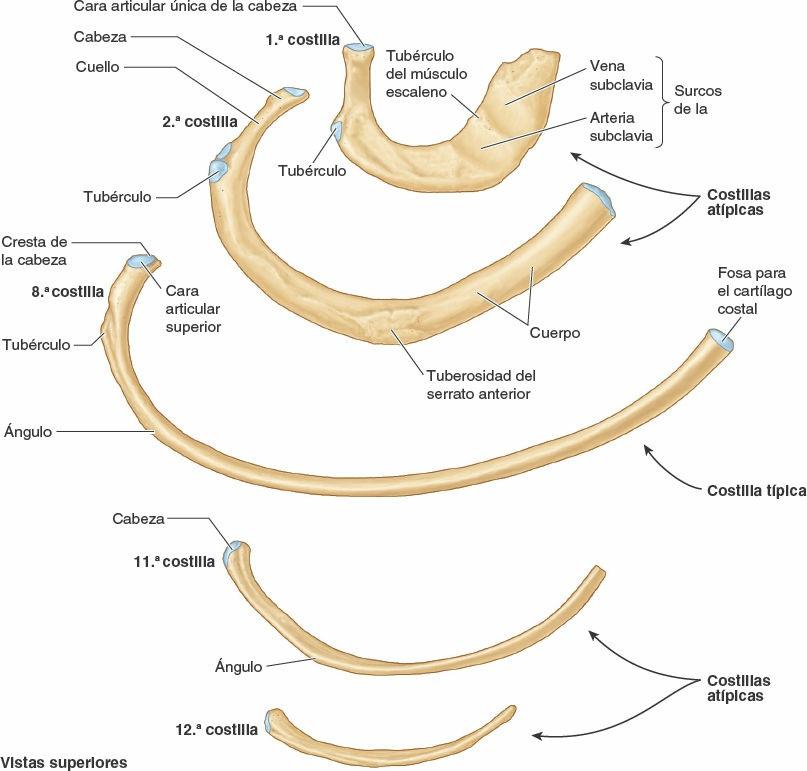
Las costillas atípicas (1ª, 2ª y 10ª-12ª) se diferencian del resto por varias características (fig. 4-4):
- La 1ª costilla es la más ancha (es decir, su cuerpo es más ancho y casi horizontal), corta y curvada de las siete costillas verdaderas. Tiene una única cara en su cabeza para articularse sólo con la vértebra T1, y dos surcos, que cruzan transversalmente su cara superior, para los vasos subclavios; los surcos están separados por una cresta y un tubérculo del músculo escaleno anterior, que proporciona inserción a dicho músculo.
- La 2ª costilla es más delgada, menos curva y más larga que la 1ª costilla. Tiene dos caras articulares en su cabeza para la articulación con los cuerpos de las vértebras T1 y T2. Su característica más atípica es un área rugosa en su cara superior, la tuberosidad del músculo serrato anterior, en la cual tiene parcialmente su origen este músculo.
- Las costillas 10ª-12ª, al igual que la 1ª costilla, tienen sólo una cara articular en sus cabezas y se articulan con una sola vértebra.
- Las costillas 11ª y 12ª son cortas y no presentan cuellos ni tubérculos.
Los cartílagos costales prolongan las costillas anteriormente y contribuyen a la elasticidad de la pared torácica, proporcionando una inserción flexible para sus terminaciones anteriores (extremos). La longitud de los cartílagos es creciente en los siete primeros, y decrece gradualmente en los siguientes. Los siete primeros cartílagos costales se insertan, de manera directa e independiente, en el esternón: el 8º, 9º y 10º se articulan con los cartílagos costales justo por encima de ellos, formando el arco costal cartilaginoso, continuo y articulado (fig. 4-1 A; también la fig. 4-13). Los cartílagos costales 11º y 12º forman bulbos sobre las extremidades anteriores de las costillas correspondientes y no llegan a ponerse en contacto ni se articulan con ningún hueso ni cartílago. Los cartílagos costales de las costillas 1ª-10ª fijan claramente el extremo anterior de la costilla al esternón, limitando su movimiento al girar su extremo posterior alrededor del eje transversal de la costilla (fig. 4-5).
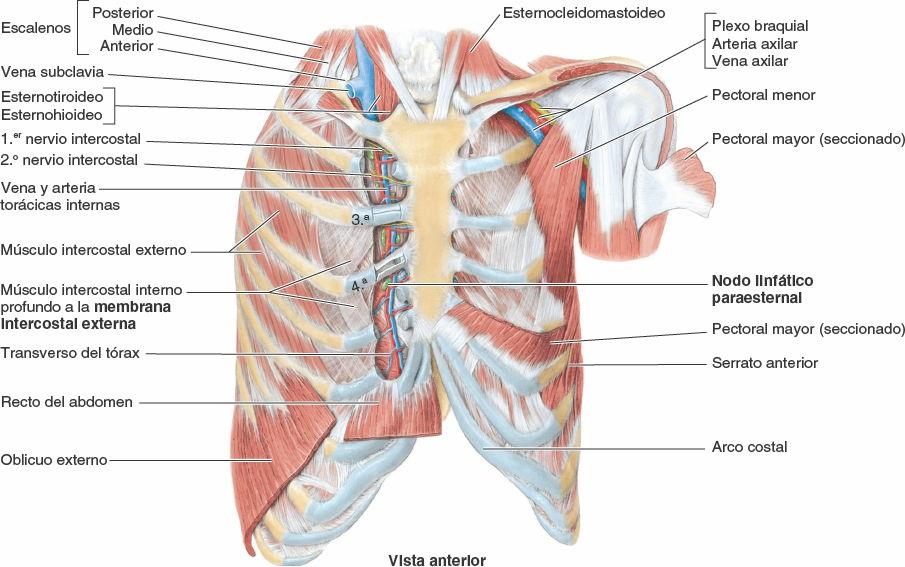
Los espacios intercostales separan las costillas y sus cartílagos costales entre sí (fig. 4-1 A). Los espacios se denominan de acuerdo con la costilla que forma el borde superior del espacio —por ejemplo, el 4º espacio intercostal está entre las costillas 4ª y 5ª. Hay 11 espacios intercostales y 11 nervios intercostales. Los espacios intercostales están ocupados por los músculos y las membranas intercostales, y dos grupos (principal y colateral) de vasos sanguíneos y nervios intercostales, que se identifican por el mismo número asignado al espacio. El espacio situado por debajo de la costilla 12ª no se encuentra entre dos costillas y por eso se denomina espacio subcostal, y el ramo anterior del nervio espinal T12 es el nervio subcostal. Los espacios intercostales son más amplios anterolateralmente, y se ensanchan todavía más durante la inspiración y con la extensión contralateral y/o flexión lateral de la columna vertebral torácica.
Vértebras torácicas
La mayoría de las vértebras torácicas son vértebras típicas, ya que son independientes y tienen cuerpo, arco vertebral y siete procesos para las uniones musculares y articulares (figs. 4-4 y 4-5). Los detalles característicos de las vértebras torácicas son:
- Dos fositas costales (hemicarillas) bilaterales en los cuerpos vertebrales, normalmente dos superiores y dos inferiores, para la articulación con las cabezas de las costillas.
- Las fositas costales en sus procesos transversos para la articulación con los tubérculos de las costillas, excepto en las dos o tres vértebras torácicas inferiores.
- Unos procesos espinosos largos inclinados inferiormente.
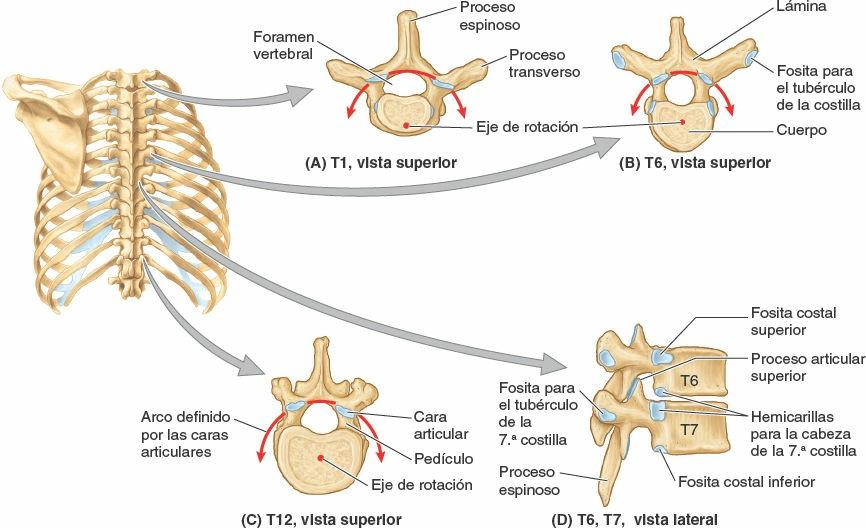
Las fositas costales superiores e inferiores, la mayoría de las cuales son pequeñas hemicarillas, se presentan emparejadas bilateralmente, como superficies planas en los bordes posterolaterales superior e inferior de los cuerpos de las vértebras torácicas típicas (T2-T9). Funcionalmente, las fositas se disponen en pares sobre las vértebras adyacentes, flanqueando un disco intervertebral interpuesto: una (hemi)carilla inferior de la vértebra superior y una (hemi)carilla superior de la vértebra inferior. Típicamente, dos hemicarillas emparejadas de esta forma y el borde posterolateral del disco intervertebral entre ellas forman una única fosa para recibir la cabeza de la costilla identificada con el mismo número que la vértebra inferior (ej. la cabeza de la 6ª costilla con la carilla costal superior de la vértebra T6). Las vértebras torácicas atípicas tienen fositas costales completas en lugar de hemicarillas:
- Las fositas costales superiores de la vértebra T1 no son hemicarillas debido a que no hay hemicarillas por encima de la vértebra C7, y la 1ª costilla se articula sólo con la vértebra T1. La T1 tiene una (hemi)carilla costal inferior típica.
- La T10 tiene un único par bilateral de fositas costales (enteras), situadas parcialmente en su cuerpo y parcialmente en su pedículo.
- Las T11 y T12 también tienen un único par de fositas costales (enteras), situadas en sus pedículos.
Los procesos espinosos se proyectan desde los arcos vertebrales de las vértebras torácicas típicas (ej. T6 o T7), son largas e inclinadas inferiormente, y normalmente se solapan con la vértebra inferior (figs. 4-4 D y 4-5). Recubren los intervalos entre las láminas de las vértebras adyacentes, evitando así que objetos cortantes como un cuchillo penetren en el conducto vertebral y lesionen la médula espinal. Las caras articulares superiores convexas de los procesos articulares superiores se orientan sobre todo posteriormente y algo lateralmente, mientras que las caras articulares inferiores cóncavas de los procesos articulares inferiores lo hacen sobre todo a nivel anterior y algo medial. Los planos articulares bilaterales entre las respectivas caras articulares de las vértebras torácicas adyacentes definen un arco, centrado en un eje de rotación dentro del cuerpo vertebral (fig. 4-4 A a C). De ese modo pueden realizarse pequeños movimientos de rotación entre las vértebras adyacentes, limitados por la parrilla costal fijada a ellas.
Esternón
El esternón es el hueso plano y alargado verticalmente que forma la parte central de la porción anterior de la caja torácica (fig. 4-6). Recubre directamente y protege las vísceras mediastínicas en general y gran parte del corazón en particular. El esternón consta de tres porciones: manubrio, cuerpo y proceso xifoides. En adolescentes y adultos jóvenes, las tres partes están unidas mediante articulaciones cartilaginosas (sincondrosis), que se osifican durante la edad adulta media o tardía.
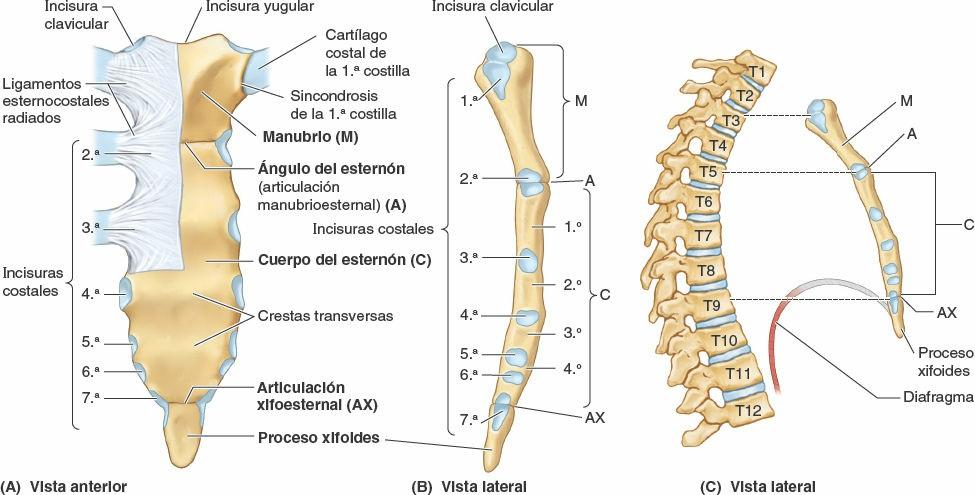
El manubrio (como la empuñadura de una espada, con el cuerpo esternal formando la hoja) es un hueso más o menos trapezoidal. El manubrio es la porción más ancha y gruesa de las tres partes del esternón. La parte central cóncava del borde superior del manubrio, la que se palpa más fácilmente, es la incisura yugular (incisura supraesternal). La incisura se acentúa por los extremos mediales (esternales) de las clavículas, las cuales son más grandes que las relativamente pequeñas incisuras claviculares del manubrio que las reciben, formando las articulaciones esternoclaviculares (fig. 4-1 A). Inferolateral a la incisura clavicular, el cartílago costal de la 1ª costilla está fuertemente unido al borde lateral del manubrio —la sincondrosis de la 1ª costilla (figs. 4-1 A y 4-6 A). El manubrio y el cuerpo del esternón se hallan en planos ligeramente diferentes superior e inferiormente a su unión, la articulación manubrioesternal (fig. 4-6 A y B); por lo tanto, su unión forma una proyección, el ángulo del esternón (de Louis).
El cuerpo del esternón es más largo, más estrecho y más delgado que el manubrio, y está localizado a nivel de las vértebras T5-T9 (fig. 4-6 A a C). Su anchura es variable debido a sus bordes laterales festoneados por las incisuras costales. En las personas jóvenes son evidentes cuatro esternebras (segmentos primordiales del esternón). Las esternebras se articulan una con otra en las articulaciones cartilaginosas primarias (sincondrosis esternales). Estas articulaciones empiezan a fusionarse desde el extremo inferior entre la pubertad (madurez sexual) y los 25 años de edad. La superficie anterior casi plana del cuerpo del esternón está marcada en los adultos por tres crestas transversas variables (fig. 4-6 A), que representan las líneas de fusión (sinostosis) de sus cuatro esternebras, originalmente separadas.
El proceso xifoides, la porción más pequeña y variable del esternón, es alargado y delgado. Su extremo inferior se encuentra al nivel de la vértebra T10. Aunque a menudo es puntiaguda, el proceso puede ser romo, bífido, curvado o estar desviado hacia un lado o anteriormente. En las personas jóvenes el proceso es cartilaginoso, pero en los adultos mayores de 40 años está más o menos osificado. En las personas ancianas, el proceso xifoides puede estar fusionado con el cuerpo del esternón.
El proceso xifoides es un importante punto de referencia del plano medio debido a que:
- Su unión con el cuerpo del esternón en la articulación xifoesternal señala el límite inferior de la parte central de la cavidad torácica; esta unión es también la localización del ángulo infraesternal (subcostal) formado por los arcos costales derecho e izquierdo (fig. 4-1 A).
- Es un marcador de la línea media para el límite superior del hígado, el centro tendinoso del diafragma y el borde inferior del corazón.
Aperturas torácicas
La caja torácica proporciona una pared periférica completa, pero está abierta superiormente e inferiormente. La apertura superior, mucho más pequeña, es un lugar de paso que permite la comunicación con el cuello y los miembros superiores. La apertura inferior, más grande, proporciona el origen del diafragma, similar a un anillo, que cierra completamente la apertura. Los desplazamientos del diafragma controlan sobre todo el volumen y la presión interna de la cavidad torácica, proporcionando la base para la respiración corriente (intercambio de aire).
Apertura torácica superior
La apertura (orificio) torácica superior está limitada (fig. 4-7):
- Posteriormente, por la vértebra T1, cuyo cuerpo protruye anteriormente en la apertura.
- Lateralmente, por el primer par de costillas y sus cartílagos costales.
- Anteriormente, por el borde superior del manubrio.
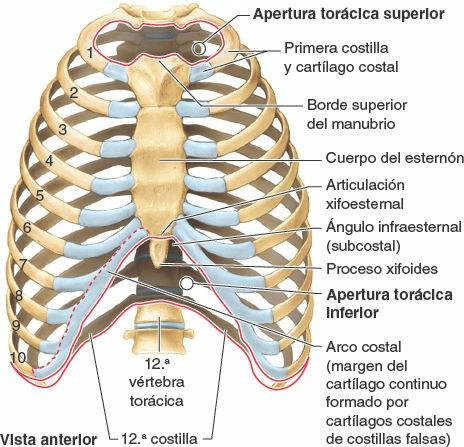
Las estructuras que pasan entre la cavidad torácica y el cuello a través de la apertura torácica superior, oblicua y en forma de riñón, son la tráquea, el esófago, los nervios y los vasos que proporcionan inervación, irrigación y drenaje a la cabeza, el cuello y los miembros superiores.
La apertura torácica superior del adulto mide unos 6,5 cm anteroposteriormente y 11 cm transversalmente. Para visualizar el tamaño de esta apertura, adviértase que es un poco más grande de lo necesario para permitir el paso de un bloque de madera de 5 × 10 cm. Debido a la oblicuidad del primer par de costillas, la apertura se inclina anteroinferiormente.
Apertura torácica inferior
La apertura (orificio) torácica inferior, la salida torácica de los anatomistas, está limitada:
- Posteriormente, por la 12ª vértebra torácica, cuyo cuerpo protruye anteriormente en la apertura.
- Posterolateralmente, por los pares de costillas 11º y 12º.
- Anterolateralmente, por la unión de los cartílagos costales de las costillas 7-10, que forman los arcos costales.
- Anteriormente, por la articulación xifoesternal.
La apertura torácica inferior es mucho más espaciosa que la apertura torácica superior y tiene un contorno irregular. También es oblicua debido a que la pared torácica posterior es mucho más larga que la anterior. Al cerrar la apertura torácica inferior, el diafragma separa las cavidades torácica y abdominal casi por completo.
Las estructuras que pasan hacia o desde el tórax al abdomen, o viceversa, lo hacen a través de las aperturas que atraviesan el diafragma (ej. el esófago y la vena cava inferior), o pasan posteriormente a este (ej. la aorta).
Al igual que a menudo se sobreestima el tamaño de la cavidad torácica (o de su contenido), su extensión inferior (que corresponde al límite entre las cavidades torácica y abdominal) también se calcula con frecuencia de forma incorrecta debido a la discrepancia entre la apertura torácica inferior y la ubicación del diafragma (suelo de la cavidad torácica) en las personas vivas. Aunque el diafragma tiene su origen en las estructuras que constituyen la apertura torácica inferior, las cúpulas del diafragma ascienden hasta el nivel del 4º espacio intercostal, y algunas vísceras abdominales, como el hígado, el bazo y el estómago, se sitúan por encima del plano de la apertura torácica inferior, dentro de la pared torácica (fig. 4-1 A y B).
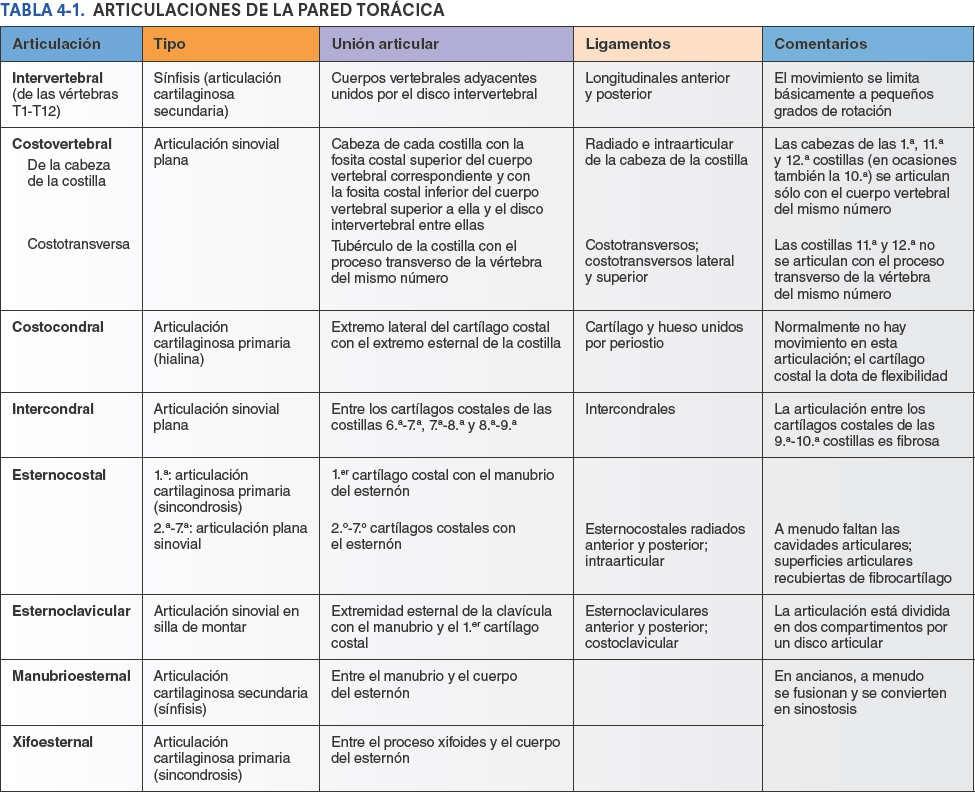
Articulaciones de la pared torácica
Aunque los movimientos de las articulaciones de la pared torácica son frecuentes —por ejemplo, asociados a la respiración normal— el grado de movimiento de cada articulación considerada individualmente es relativamente pequeño. No obstante, cualquier alteración que reduzca la movilidad de estas articulaciones interfiere con la respiración. Durante la respiración profunda, los movimientos de la caja torácica (anterior, superior o lateralmente) son considerables. Al extender la columna vertebral se aumenta adicionalmente el diámetro anteroposterior del tórax. Las articulaciones de la pared torácica se ilustran en la figura 4-8. El tipo, las superficies articulares implicadas y los ligamentos de las articulaciones de la pared torácica se detallan en la tabla 4-1.
Las articulaciones intervertebrales entre los cuerpos de las vértebras adyacentes están unidas por ligamentos longitudinales y por discos intervertebrales.
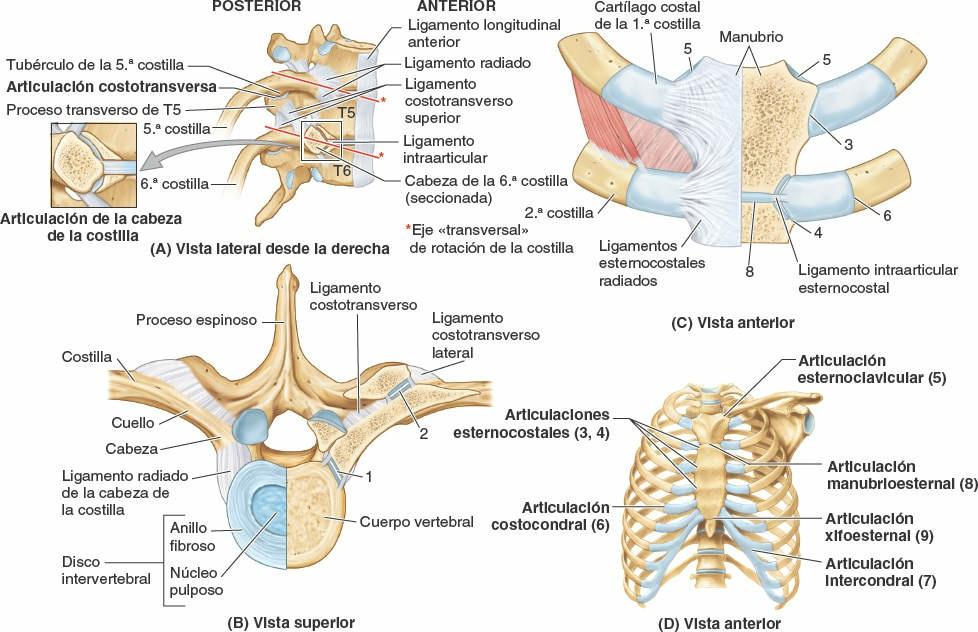
Articulaciones costovertebrales
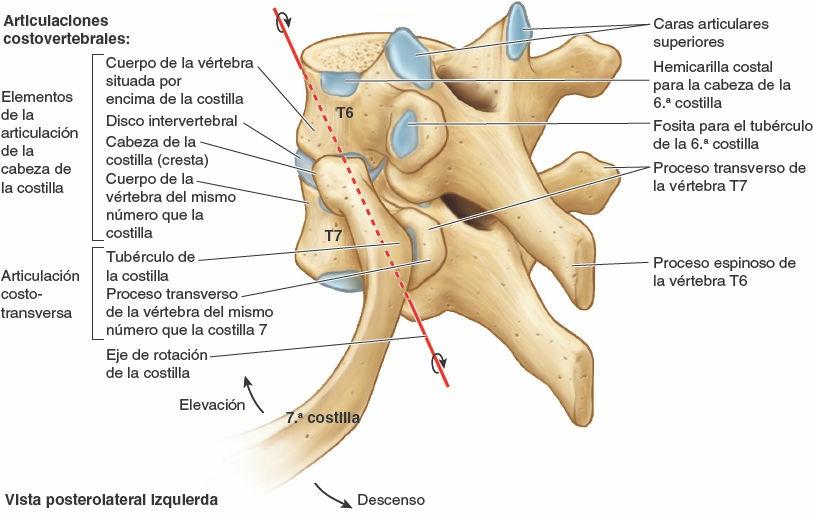
Una costilla típica se articula posteriormente con la columna vertebral mediante dos articulaciones: la articulación de la cabeza de la costilla y la articulación costotransversa (fig. 4-5).
Articulaciones de las cabezas de las costillas
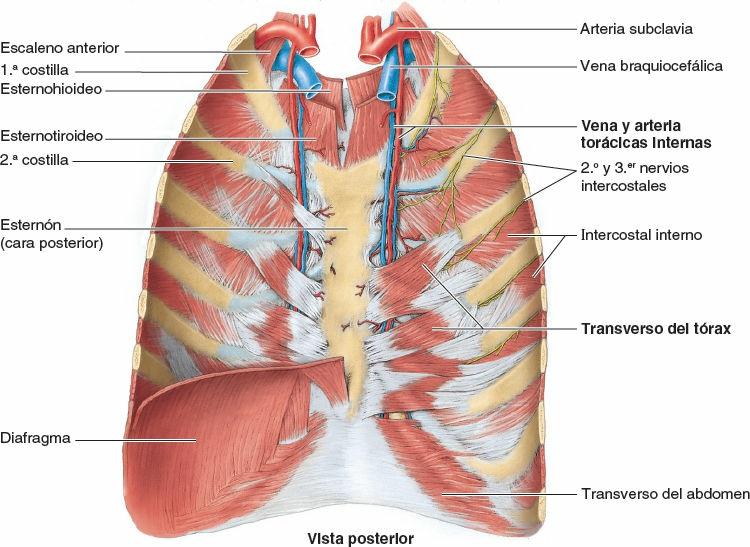
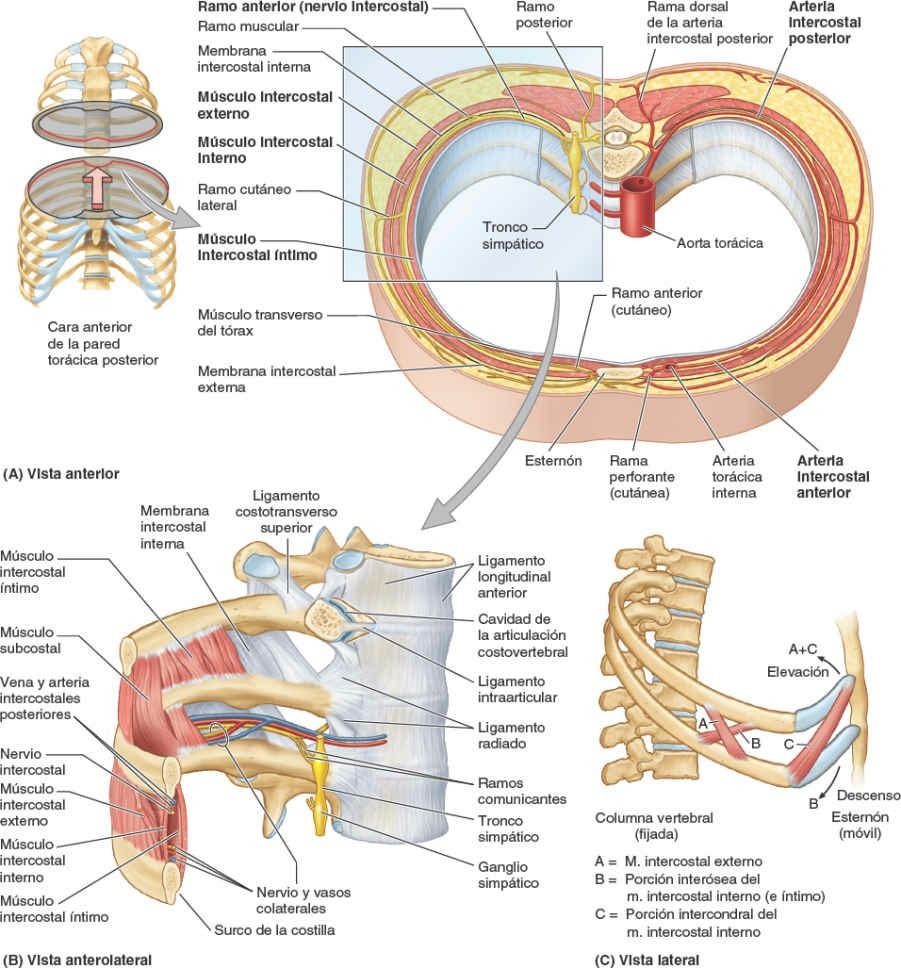
La cabeza de la costilla se articula con la fosita costal superior de la vértebra correspondiente (del mismo número), con la fosita costal inferior de la vértebra superior a ella, y con el disco intervertebral adyacente que une las dos vértebras (figs. 4-4 y 4-8 A). Por ejemplo, la cabeza de la 6ª costilla se articula con la fosita costal superior del cuerpo de la vértebra T6, con la fosita costal inferior de la T5 y con el disco intervertebral situado entre estas dos vértebras. La cresta de la cabeza de la costilla se une al disco intervertebral por un ligamento intraarticular de la cabeza de la costilla dentro de la articulación, dividiendo este espacio cerrado en dos cavidades sinoviales.
La membrana fibrosa de la cápsula articular es más fuerte anteriormente, donde forma un ligamento radiado de la cabeza de la costilla que se abre en abanico desde el borde anterior de la cabeza de la costilla hacia los lados de los cuerpos de dos vértebras y el disco intervertebral que hay entre ellas (fig. 4-8 A y B). Las cabezas de las costillas se unen tan estrechamente a los cuerpos vertebrales que sólo permiten leves movimientos de deslizamiento en las (hemi)carillas (girando alrededor del ligamento intraarticular de las articulaciones de las cabezas de las costillas). Sin embargo, incluso estos leves movimientos pueden producir un desplazamiento relativamente grande del extremo distal (esternal o anterior) de una costilla.
Articulaciones costotransversas
Existen abundantes ligamentos laterales a las porciones posteriores (arcos vertebrales) de las vértebras que refuerzan estas articulaciones y limitan sus movimientos, pues sólo poseen unas finas cápsulas articulares. El ligamento costotransverso, que pasa desde el cuello de la costilla hasta el proceso transverso, y el ligamento costotransverso lateral, que pasa desde el tubérculo de la costilla hasta el vértice del proceso transverso, fortalecen las caras anterior y posterior de la articulación, respectivamente. El ligamento costotransverso superior es una banda ancha que une la cresta del cuello de la costilla al proceso transverso superior a ella. La apertura que se forma entre este ligamento y la vértebra permite el paso del nervio espinal y de la rama posterior de la arteria intercostal. El ligamento costotransverso superior puede dividirse en un ligamento costotransverso anterior fuerte y un ligamento costotransverso posterior débil.
Los fuertes ligamentos costotransversos que unen estas articulaciones limitan sus movimientos a un leve deslizamiento. Sin embargo, las superficies articulares de los tubérculos de las seis costillas superiores son convexos y encajan en las concavidades de los procesos transversos (fig. 4-9). Como resultado, se produce la rotación alrededor de un eje transversal en su mayor parte que atraviesa el ligamento intraarticular y la cabeza y el cuello de la costilla (fig. 4-8 A y B). Esto produce movimientos de elevación y descenso de los extremos esternales de las costillas y del esternón en un plano sagital (movimiento de palanca de bomba de agua) (fig. 4-10 A y C). Las superficies articulares planas de los tubérculos y los procesos transversos de las costillas 7ª-10ª permiten el deslizamiento (fig. 4-9), produciendo la elevación y el descenso de las porciones más laterales de estas costillas en un plano transversal (movimiento de asa de cubo) (fig. 4-10 B y C).
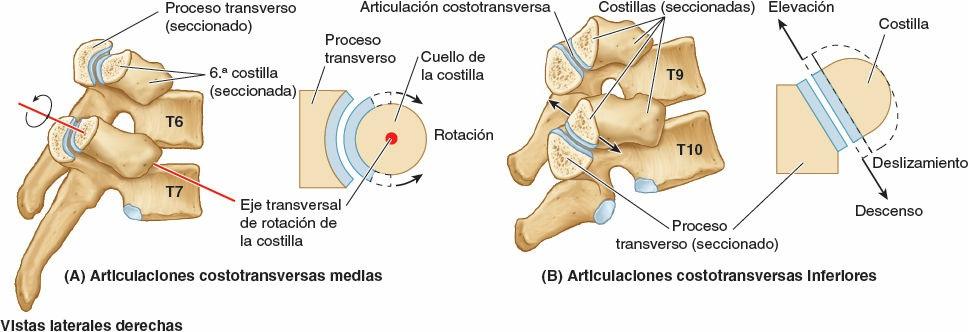
Articulaciones esternocostales
El primer par de cartílagos costales se articula con el manubrio por medio de una fina y densa lámina de fibrocartílago fuertemente adherido interpuesto entre el cartílago y el manubrio, la sincondrosis de la 1ª costilla. Los pares 2º-7º de cartílagos costales se articulan con el esternón por medio de articulaciones sinoviales, con superficies articulares fibrocartilaginosas en ambas caras, condral y esternal, que permiten el movimiento durante la respiración. Las débiles cápsulas articulares de estas articulaciones se espesan anterior y posteriormente para formar los ligamentos esternocostales radiados. Estos continúan como bandas finas y anchas que pasan desde los cartílagos costales hasta las superficies anterior y posterior del esternón, formando una cubierta, parecida a un fieltro, para este hueso.
Movimientos de la pared torácica
Los movimientos de la pared torácica y del diafragma durante la inspiración aumentan el volumen intratorácico y los diámetros del tórax (fig. 4-10 D y F). En consecuencia, los cambios de presión provocan alternativamente la aspiración de aire hacia el interior de los pulmones (inspiración), a través de la nariz, la boca, la laringe y la tráquea, y su expulsión de los pulmones (espiración) a través de los mismos conductos. Durante la espiración pasiva, el diafragma, los músculos intercostales y otros, se relajan, con lo que disminuye el volumen intratorácico y aumenta la presión intratorácica (fig. 4-10 E y C). Al mismo tiempo, la presión intraabdominal disminuye y las vísceras abdominales se descomprimen. El tejido elástico de los pulmones, que estaba expandido, se retrae, y se expele la mayor parte del aire.
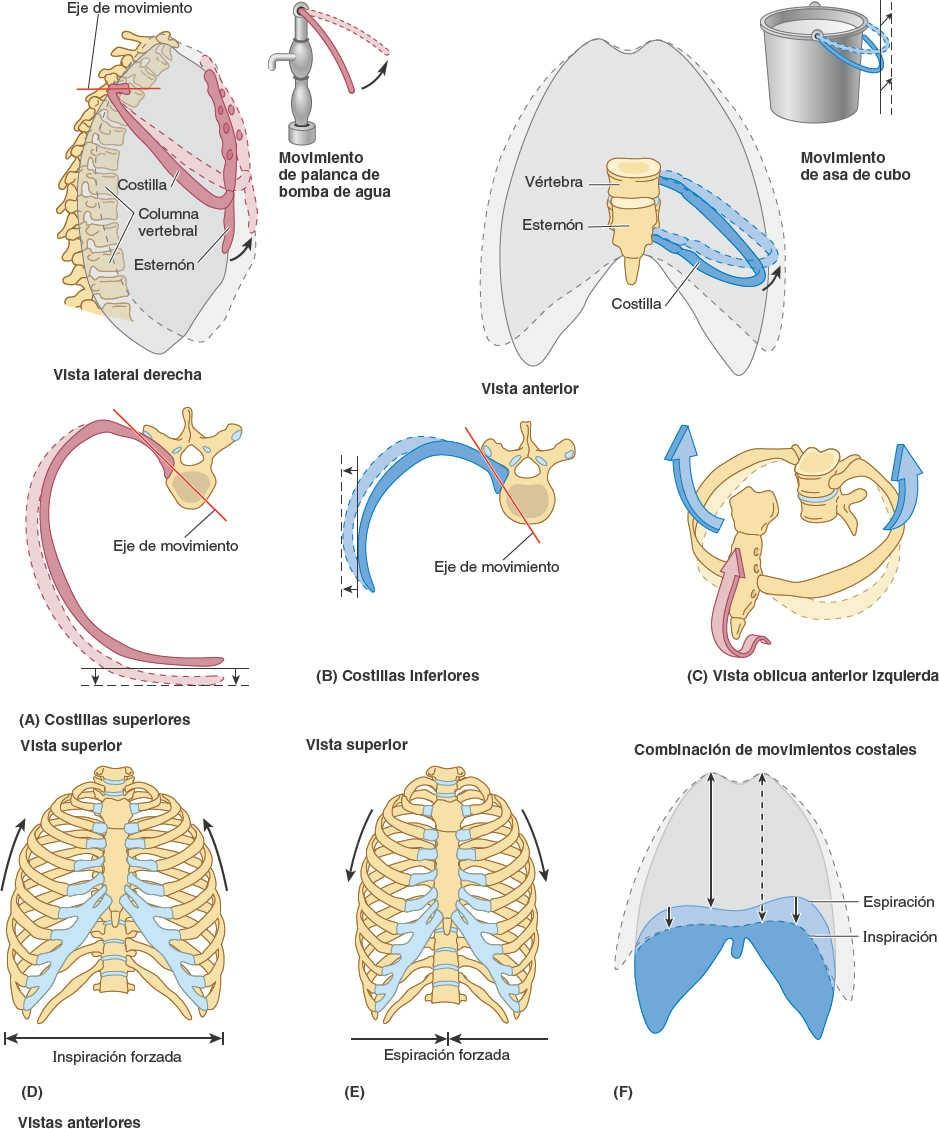
El diámetro vertical (altura) de la parte central de la cavidad torácica aumenta durante la inspiración a medida que el diafragma desciende al contraerse, comprimiendo las vísceras abdominales (fig. 4-10 F). Durante la espiración, el diámetro vertical vuelve a su posición neutra a medida que la retracción de los pulmones genera una presión subatmosférica en las cavidades pleurales, entre los pulmones y la pared torácica. Como resultado de ello y de la ausencia de resistencia sobre las vísceras previamente comprimidas, las cúpulas del diafragma ascienden y se reduce el diámetro vertical.
El diámetro anteroposterior del tórax aumenta considerablemente cuando se contraen los músculos intercostales. El movimiento de las costillas (fundamentalmente 2ª-6ª) en las articulaciones costovertebrales, alrededor de un eje que pasa a través del cuello de las costillas, hace que los extremos anteriores de las costillas asciendan —el movimiento de palanca de bomba de agua (fig. 4-10 A y C). Debido a que las costillas se inclinan inferiormente, su elevación provoca también un movimiento anteroposterior del esternón, en especial de su extremo inferior, con un ligero movimiento que en las personas jóvenes tiene lugar en la articulación manubrioesternal, cuando la articulación aún no se ha fusionado (unido).
El diámetro transverso del tórax también aumenta ligeramente cuando se contraen los músculos intercostales, elevando las partes medias (las más laterales) de las costillas (en especial las más inferiores) —el movimiento de asa de cubo (fig. 4-10 B y C). La combinación de todos estos movimientos mueve la caja torácica anterior, superior y lateralmente (fig. 4-10 C y F).
Músculos de la pared torácica
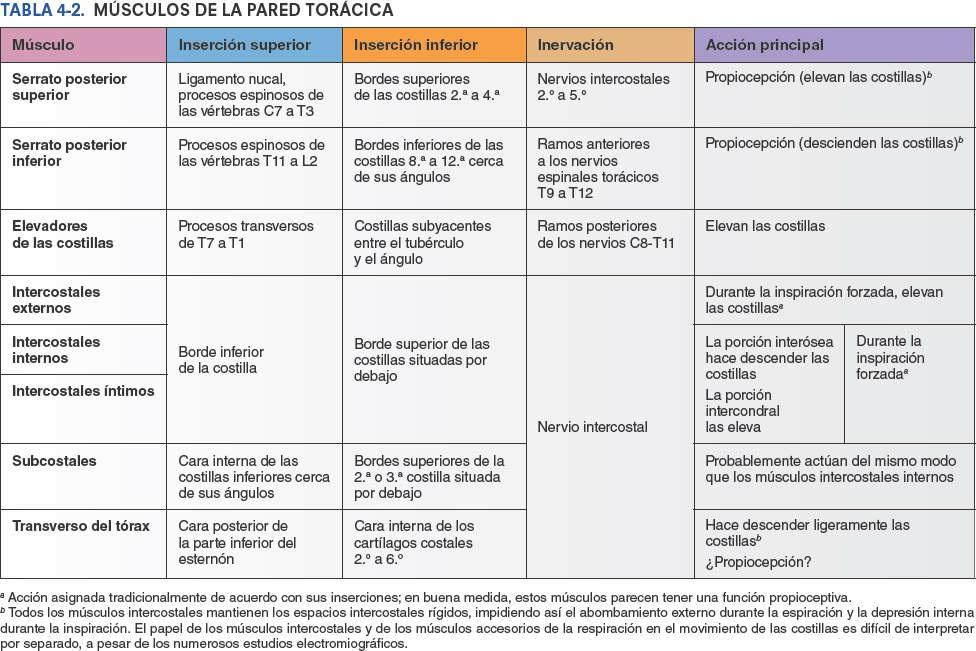
Algunos músculos que están insertados en la caja torácica, o que la cubren, están implicados fundamentalmente en acciones sobre otras regiones anatómicas. Los músculos axioapendiculares se extienden desde la caja torácica (esqueleto axial) hasta los huesos del miembro superior (esqueleto apendicular). De forma similar, algunos músculos de la pared anterolateral del abdomen, el dorso y el cuello tienen su inserción en la caja torácica (fig. 4-11). Los músculos axioapendiculares actúan sobre todo en los miembros superiores, pero varios de ellos, como el pectoral mayor, el pectoral menor y la porción inferior del serrato anterior, también pueden ejercer como músculos accesorios de la respiración, ayudando a elevar las costillas para expandir la cavidad torácica cuando la inspiración es profunda y forzada (ej. tras correr los 100 m lisos). Los músculos escalenos, que descienden desde las vértebras del cuello hasta la 1ª y la 2ª costillas, actúan principalmente sobre la columna vertebral. Sin embargo, también actúan como músculos accesorios de la respiración fijando estas costillas y permitiendo que los músculos que unen las costillas situadas por debajo sean más eficaces para elevar las costillas inferiores durante la inspiración forzada.
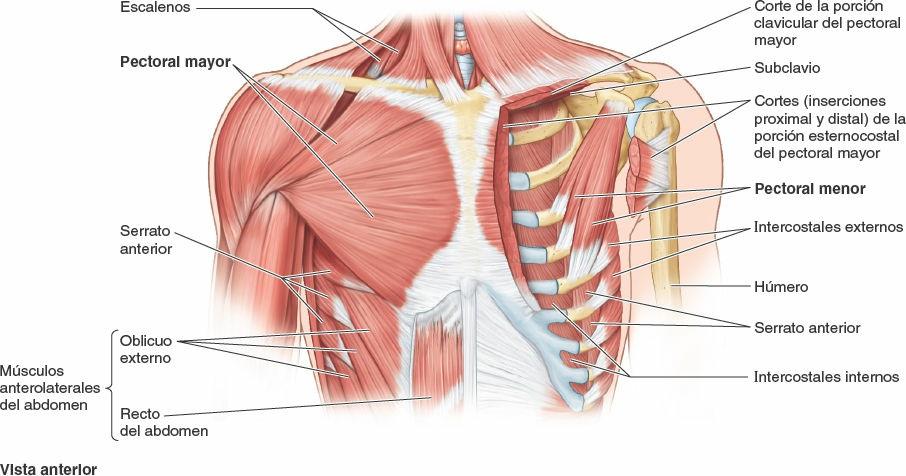
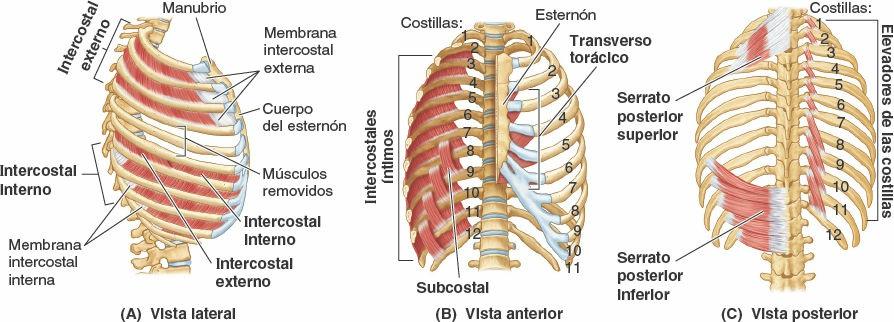
Los verdaderos músculos de la pared torácica son los serratos posteriores, los elevadores de las costillas, los intercostales, los subcostales y los transversos del tórax; se muestran en la figura 4-12 A y B, y sus inserciones, inervación y funciones se detallan en la tabla 4-2.
Los músculos serratos posteriores se han considerado tradicionalmente músculos inspiratorios, aunque esta función no está respaldada por la electromiografía ni por cualquier otra evidencia. Según sus inserciones y disposición, el serrato posterior superior se decía que elevaba las cuatro costillas superiores, aumentando el diámetro anteroposterior del tórax y elevando el esternón. Según sus inserciones y disposición, se afirmaba que el serrato posterior inferior descendía las costillas inferiores, evitando que el diafragma las arrastrase hacia arriba. Pero, estudios recientes (Vilensky et al., 2001) sugieren que estos músculos, que se extienden sobre las aperturas torácicas superior e inferior, así como las transiciones desde la columna vertebral torácica relativamente inflexible hasta los segmentos cervical y lumbar de la columna, mucho más flexibles, podrían no tener una función principalmente motora. En lugar de eso, podrían tener una función propioceptiva. Estos músculos, en particular el serrato posterior superior, han sido implicados como fuente de dolor crónico en las fibromialgias.
Los músculos elevadores de las costillas son 12 músculos en forma de abanico que elevan las costillas (fig. 4-17), aunque su papel en la inspiración normal, si tienen alguno, no está claro. Pueden participar en el movimiento vertebral y/o la propiocepción.
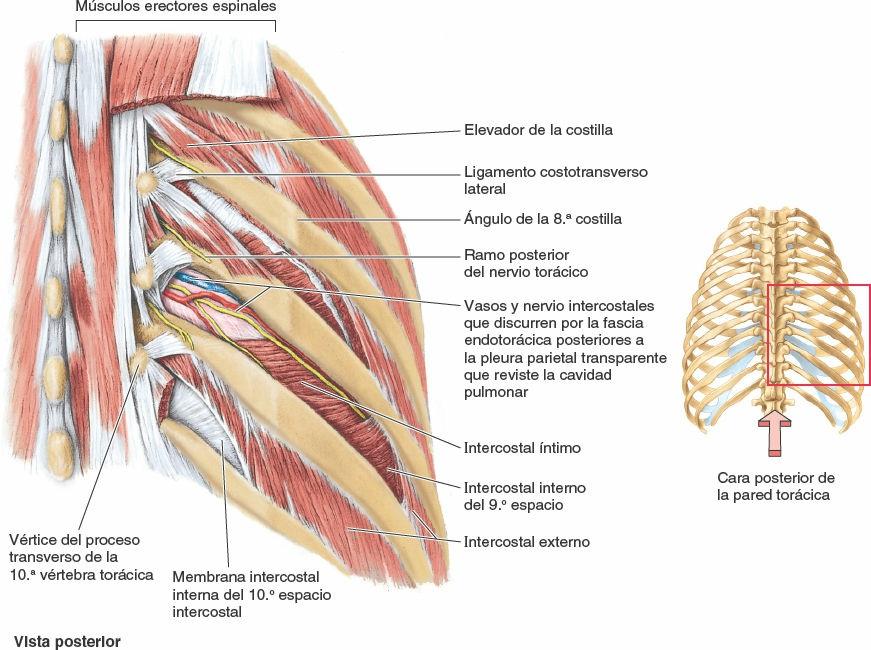
Los músculos intercostales ocupan los espacios intercostales (figs. 4-11 a 4-14; tabla 4-2). La capa superficial está formada por los intercostales externos, y la capa interna por los intercostales internos. A las fibras más profundas de estos últimos, dispuestas internamente en los vasos intercostales, se les designa, un tanto artificialmente, como un músculo diferenciado, los intercostales íntimos.
- Los músculos intercostales externos (11 pares) ocupan los espacios intercostales desde los tubérculos de las costillas posteriormente hasta las uniones costocondrales anteriormente (figs. 4-11 a 4-13, y 4-15). Anteriormente, las fibras musculares son reemplazadas por las membranas intercostales externas (fig. 4-15 A). Estos músculos discurren a nivel inferoanterior desde la costilla superior hasta la inferior. Cada músculo se inserta superiormente en el borde inferior de la costilla que tiene por encima, e inferiormente en el borde superior de la costilla que tiene por debajo (fig. 4-15 C). Estos músculos se continúan inferiormente con los músculos oblicuos externos en la pared anterolateral del abdomen. Los intercostales externos son más activos durante la inspiración.
- Los músculos intercostales internos (11 pares) discurren profunda y perpendicularmente a los intercostales externos (figs. 4-12 B, 4-14 y 4-15 C). Sus fibras discurren a nivel inferoposterior desde el suelo de los surcos costales hasta los bordes superiores de las costillas inferiores a ellos. Los intercostales internos se insertan en los cuerpos de las costillas y a sus cartílagos costales, alcanzando anteriormente el esternón y posteriormente el ángulo de las costillas (fig. 4-16). Entre las costillas posteriormente, medial a los ángulos, los intercostales internos son sustituidos por las membranas intercostales internas (fig. 4-15 A). Los músculos intercostales internos inferiores se continúan con los músculos oblicuos internos en la pared anterolateral del abdomen. Los intercostales internos —más débiles que los músculos intercostales externos— son más activos durante la espiración, especialmente sus porciones interóseas (frente a las intercondrales).
- Los músculos intercostales íntimos son similares a los intercostales internos y son, en esencia, sus porciones más profundas. Los intercostales íntimos están separados de los intercostales internos por los vasos y los nervios intercostales (figs. 4-15 A y B, y 4-16). Estos músculos se extienden entre las superficies internas de las costillas adyacentes y ocupan las partes más laterales de los espacios intercostales. Es probable (aunque no está establecido) que sus acciones sean las mismas que las de los músculos intercostales internos.
Los músculos subcostales tienen formas y tamaños variables, y normalmente sólo están bien desarrollados en la pared torácica inferior. Estas finas tiras musculares se extienden desde la superficie interna del ángulo de una costilla hasta la superficie interna de la segunda o tercera costilla inferior a ella. Atravesando uno o dos espacios intercostales, los subcostales discurren en la misma dirección que los intercostales internos y se mezclan con ellos (fig. 4-15 B).
Los músculos transversos del tórax constan de cuatro o cinco tiras que irradian superolateralmente desde la cara posterior de la porción inferior del esternón (figs. 4-13 a 4-15 A). Los músculos transversos del tórax se continúan inferiormente con los músculos transversos del abdomen en la pared anterolateral del cuerpo. Estos músculos parecen tener una débil función espiratoria y también pueden proporcionar información propioceptiva.
Aunque los intercostales externos e internos son activos durante la inspiración y la espiración, respectivamente, la mayor parte de su actividad es isométrica (aumento de tono sin producir movimiento); el papel de estos músculos en el movimiento de las costillas parece tener relación, sobre todo, con la respiración forzada. El diafragma es el principal músculo de la inspiración. La espiración es pasiva a no ser que se exhale en contra de una resistencia (ej. al inflar un globo) o cuando se intenta exhalar el aire más rápidamente de lo habitual (ej. al toser, estornudar, sonarse la nariz o gritar). La retracción elástica de los pulmones y la descompresión de las vísceras abdominales expelen el aire inhalado previamente.
El papel primordial de los músculos intercostales en la respiración es sostener (aumentar el tono o la rigidez) el espacio intercostal, oponiéndose al movimiento paradójico, en especial durante la inspiración, cuando las presiones torácicas internas son más bajas (más negativas). Esto se hace más evidente después de una lesión medular alta, cuando inicialmente hay una parálisis fláccida de todo el tronco aunque el diafragma se mantiene activo. En estas circunstancias, la capacidad vital está seriamente afectada por la incursión paradójica de la pared torácica durante la inspiración. Varias semanas después, la parálisis se hace espástica; la pared torácica se vuelve más rígida y aumenta la capacidad vital.
La acción mecánica de los músculos intercostales en el movimiento de la costilla, en especial durante la respiración forzada, se puede apreciar mediante un modelo simple (fig. 4-15 C). Un par de palancas curvas, que representan las costillas que limitan un espacio intercostal, se unen posteriormente a una columna vertebral fija y anteriormente a un esternón móvil. Las costillas (y el espacio intercostal interpuesto) descienden a medida que se desplazan anteriormente, alcanzando su punto más bajo aproximadamente en la unión costocondral, y luego ascienden hasta el esternón. Los músculos cuyas fibras tienen una inclinación más parecida a la de las costillas en sus inserciones (intercostales externos y la porción intercondral de los músculos intercostales internos) rotan superiormente las costillas en sus ejes posteriores, elevando las costillas y el esternón. Los músculos con fibras aproximadamente perpendiculares a la inclinación de las costillas en sus inserciones (porción interósea de los músculos intercostales internos) rotan inferiormente las costillas en sus ejes posteriores, descendiendo las costillas y el esternón.
El diafragma es una pared compartida (en realidad, un suelo/techo) que separa el tórax del abdomen. Aunque tiene funciones relacionadas con ambos compartimentos del tronco, su función más importante (vital) es la de ser el músculo principal de la inspiración. La descripción detallada del diafragma torácico aparece en el capítulo dedicado al Abdomen, debido a que las inserciones de sus pilares están en niveles abdominales (esto es, en las vértebras lumbares) y todas sus inserciones se observan mejor desde su cara inferior (abdominal).
Fascia de la pared torácica
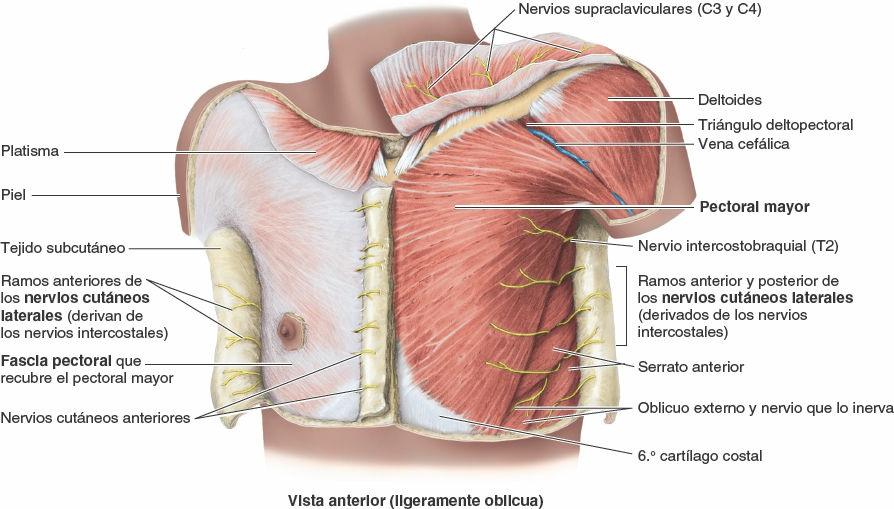
Cada parte de la fascia profunda se denomina según el músculo que reviste o la(s) estructura(s) a la(s) cual(es) está unida. Por consiguiente, una gran porción de la fascia profunda que recubre la pared torácica anterior se denomina fascia pectoral por su relación con los músculos pectorales mayores (fig. 4-16). Una buena porción de la fascia pectoral forma la mayor parte del lecho de la mama (estructuras sobre las cuales descansa la cara posterior de la mama). Profunda al pectoral mayor y a su fascia hay otra lámina de fascia profunda que, suspendida desde la clavícula, envuelve el músculo pectoral menor, la fascia clavipectoral.
La caja torácica está tapizada internamente por la fascia endotorácica (fig. 4-30 C). Esta fina lámina fibroareolar fija a la pared torácica la porción adyacente del revestimiento de las cavidades del pulmón (la pleura parietal costal). Se vuelve más fibrosa a nivel de los vértices pulmonares (membrana suprapleural).
Nervios de la cavidad torácica
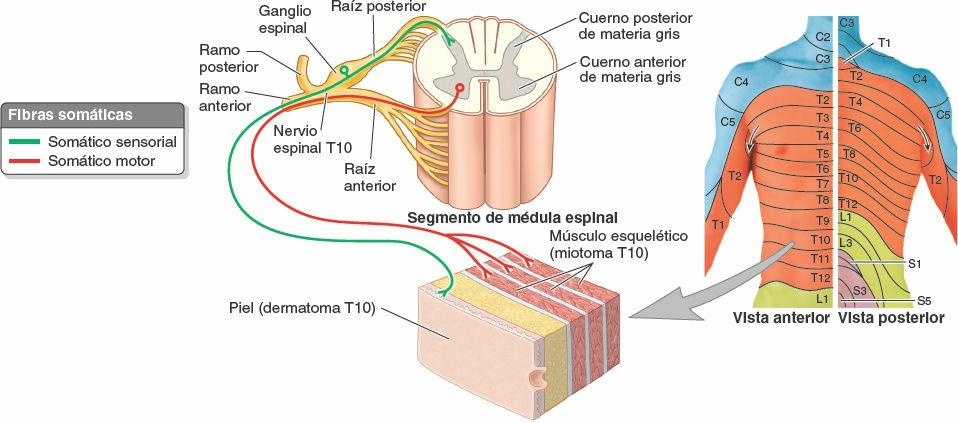
La pared torácica tiene 12 pares de nervios espinales torácicos. En cuanto salen de los forámenes (orificios) intervertebrales, se dividen en ramos anterior y posterior (ramos primarios) (figs. 4-15 A y 4-17). Los ramos anteriores de T1-T11 forman los nervios intercostales que discurren a lo largo de los espacios intercostales. El ramo anterior del nervio T12, inferior a la 12ª costilla, forma el nervio subcostal. Los ramos posteriores de los nervios espinales torácicos se dirigen hacia atrás, inmediatamente laterales a los procesos articulares de las vértebras, para inervar las articulaciones, los músculos profundos del dorso y la piel del dorso en la región torácica.
Nervios intercostales típicos
Los nervios intercostales 3º-6º entran en las porciones más mediales de los espacios intercostales posteriores; inicialmente discurren dentro de la fascia endotorácica, entre la pleura parietal (revestimiento seroso de la cavidad torácica) y la membrana intercostal interna cerca de la línea media de los espacios intercostales (figs. 4-15 A y B, y 4-17). Cerca de los ángulos de las costillas, los nervios pasan entre los músculos intercostales internos e íntimos. Ahí, los nervios intercostales entran en los surcos de las costillas y luego continúan su recorrido por ellos, situándose en o justo inferiores a los surcos de las costillas, y discurriendo inferiores a las arterias intercostales (que a su vez son inferiores a las venas intercostales). Por tanto, los paquetes neurovasculares (y especialmente los vasos) están protegidos por los bordes inferiores de las costillas suprayacentes. Los ramos colaterales de estos nervios se originan cerca de los ángulos de las costillas y discurren a lo largo del borde superior de la costilla situada inferiormente. Los nervios continúan anteriormente entre los músculos intercostales internos e íntimos, proporcionan ramos para estos y otros músculos, y dan origen a ramos cutáneos laterales, aproximadamente en la línea axilar media. Anteriormente, los nervios se encuentran sobre la cara interna del músculo intercostal interno. Próximos al esternón, los nervios giran anteriormente, pasando entre los cartílagos costales, y entran en el tejido subcutáneo como ramos cutáneos anteriores.
Mediante su ramo posterior y los ramos cutáneos lateral y anterior de su ramo anterior, la mayoría de los nervios espinales torácicos (T2-T12) inervan un área del tronco semejante a una franja, o dermatoma, que se extiende desde la línea media posterior a la línea media anterior (fig. 4-18). El grupo de músculos inervados por los ramos anterior y posterior (nervio intercostal) de cada par de nervios espinales torácicos constituye un miotoma. Los miotomas de la mayoría de los nervios espinales torácicos (T2-T11) incluyen los músculos intercostales, subcostales, transversos del tórax, elevadores de las costillas y serratos posteriores relacionados con el espacio intercostal que incluye el ramo anterior (nervio intercostal) del nervio espinal específico, más la porción suprayacente de los músculos profundos del dorso.
Los ramos de un nervio intercostal típico son los siguientes (fig. 4-15 A y B):
- Ramos comunicantes, que conectan cada nervio intercostal al tronco simpático homolateral. Las fibras presinápticas abandonan las porciones iniciales del ramo anterior de cada nervio torácico (y lumbar superior) mediante un ramo comunicante blanco y pasan hacia un tronco simpático. Las fibras postsinápticas que se distribuyen por la pared corporal y los miembros pasan desde los ganglios del tronco simpático, a través de los ramos comunicantes grises, para unirse al ramo anterior del nervio espinal más cercano, incluidos todos los nervios intercostales. Las fibras nerviosas simpáticas se distribuyen a través de todos los ramos de todos los nervios espinales (ramos anteriores y posteriores) para alcanzar los vasos sanguíneos, las glándulas sudoríparas y la musculatura lisa de la pared corporal y de los miembros.
- Ramos colaterales, que se originan cerca de los ángulos de las costillas y descienden para correr por el borde superior de la costilla inferior, colaborando en la inervación de los músculos intercostales y la pleura parietal.
- Ramos cutáneos laterales, que se originan cerca de la línea axilar media, atraviesan los músculos intercostales internos y externos y se dividen a su vez en ramos anterior y posterior. Estos ramos terminales inervan la piel de las paredes laterales torácica y abdominal.
- Ramos cutáneos anteriores, que atraviesan los músculos y las membranas del espacio intercostal en la línea paraesternal y se dividen en ramos medial y lateral. Estos ramos terminales inervan la piel de la cara anterior del tórax y el abdomen.
- Ramos musculares, que inervan los músculos intercostales, subcostales, transversos del tórax, elevadores de las costillas y serratos posteriores.
Nervios intercostales atípicos
Aunque el ramo anterior de la mayoría de los nervios espinales torácicos es simplemente el nervio intercostal para ese nivel, el ramo anterior del primer nervio espinal torácico (T1) se divide primero en una porción superior grande y una inferior pequeña. La porción superior se une al plexo braquial, el plexo nervioso que inerva el miembro superior, y la porción inferior se convierte en el primer nervio intercostal. Otras características atípicas de algunos nervios intercostales específicos son las siguientes:
- El primer y el 2º nervios intercostales recorren la cara interna de las costillas 1ª y 2ª, en vez de hacerlo a lo largo del borde inferior de los surcos de las costillas (fig. 4-14).
- El primer nervio intercostal no tiene ramo cutáneo anterior, y a menudo tampoco ramo cutáneo lateral. Cuando existe un ramo cutáneo lateral, inerva la piel de la axila y puede comunicar tanto con el nervio intercostobraquial como con el nervio cutáneo medial del brazo.
- El 2º (y a veces el tercer) nervio intercostal da origen a un gran ramo cutáneo lateral, el nervio intercostobraquial, que surge del 2º espacio intercostal en la línea axilar media, penetra el serrato anterior y entra en la axila y el brazo. Normalmente, el nervio intercostobraquial inerva el suelo—piel y tejido subcutáneo— de la axila y luego comunica con el nervio cutáneo medial del brazo para inervar las caras medial y posterior del brazo. El ramo cutáneo lateral del tercer nervio intercostal frecuentemente da origen a un segundo nervio intercostobraquial.
- Los nervios intercostales 7º-11º, tras dar origen a ramos cutáneos laterales, atraviesan posteriormente el arco costal y continúan para inervar la piel y los músculos abdominales. Cuando abandonan el espacio intercostal se convierten en nervios toracoabdominales de la pared anterior del abdomen. Sus ramos cutáneos anteriores perforan la vaina del recto, convirtiéndose en cutáneos en la proximidad del plano medio.
Vascularización de la pared torácica
En líneas generales, el patrón de la distribución vascular en la pared torácica refleja la estructura de la caja torácica —esto es, discurre por los espacios intercostales en paralelo a las costillas.
Arterias de la pared torácica
La irrigación arterial de la pared torácica (fig. 4-19; tabla 4-3) deriva de:
- La aorta torácica, a través de las arterias intercostales posteriores y subcostal.
- La arteria subclavia, a través de las arterias torácica interna e intercostal suprema.
- La arteria axilar, a través de las arterias torácicas superior y lateral.
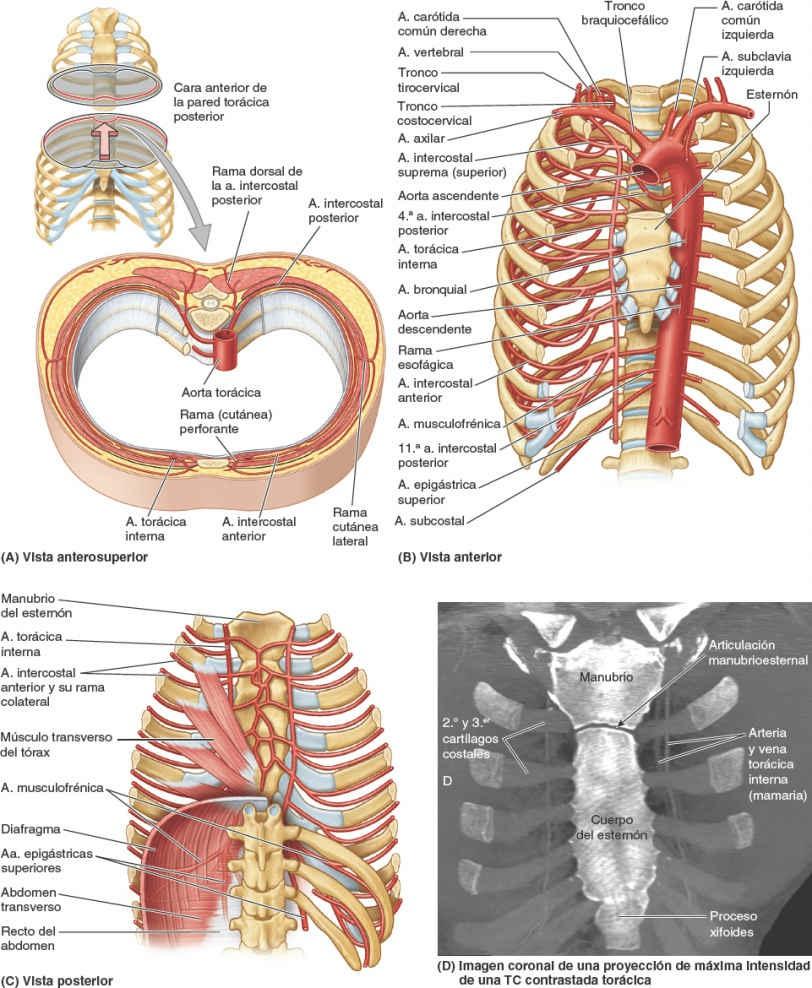
Las arterias intercostales discurren por la pared torácica entre las costillas. Con la excepción de los espacios intercostales 10º y 11º, cada espacio intercostal es irrigado por tres arterias: una gran arteria intercostal posterior (y su rama colateral) y un par de pequeñas arterias intercostales anteriores.
Las arterias intercostales posteriores:
- Del primer y 2º espacios intercostales se originan de la arteria intercostal suprema (superior), una rama del tronco costocervical de la arteria subclavia.
- De los espacios intercostales 3º-11º (y las arterias subcostales del espacio subcostal) se originan posteriormente de la aorta torácica (fig. 4-19). Debido a que la aorta está ligeramente a la izquierda de la columna vertebral, las arterias intercostales derechas 3ª-11ª cruzan sobre los cuerpos vertebrales, siguiendo un curso más largo que las del lado izquierdo (fig. 4-19 B).
- Todas emiten una rama posterior que acompaña al ramo posterior del nervio espinal para irrigar la médula espinal, la columna vertebral, los músculos del dorso y la piel.
- Dan lugar a una pequeña rama colateral que cruza el espacio intercostal y discurre a lo largo del borde superior de la costilla.
- Acompañan a los nervios intercostales a través de los espacios intercostales. Cerca del ángulo de la costilla, las arterias entran en los surcos de las costillas, donde se sitúan entre la vena y el nervio intercostales. Primero, las arterias discurren en la fascia endotorácica entre la pleura parietal y la membrana intercostal interna (fig. 4-17); después discurren entre los músculos intercostales íntimos e internos.
- Tienen ramas terminales y colaterales que se anastomosan anteriormente con las arterias intercostales anteriores (fig. 4-19 A).
Las arterias torácicas internas (históricamente, arterias mamarias internas):
- Se originan en la raíz del cuello, en las caras inferiores de las primeras porciones de las arterias subclavias.
- Descienden hacia el interior del tórax posteriormente a la clavícula y al primer cartílago costal (figs. 4-13, 4-14 y 4-19).
- Se cruzan cerca de su origen con el nervio frénico homolateral.
- Descienden sobre la cara interna del tórax ligeramente laterales al esternón y posteriores a los sextos cartílagos costales superiores y los músculos intercostales internos interpuestos. Después de descender más allá del 2º cartílago costal, las arterias torácicas internas discurren anteriores al músculo transverso del tórax (figs. 4-15 A y 4-19 C). Entre las tiras del músculo, las arterias entran en contacto con la pleura parietal posteriormente.
- Terminan en el 6º espacio intercostal dividiéndose en las arterias epigástrica superior y musculofrénica.
- Dan lugar directamente a las arterias intercostales anteriores que irrigan los seis espacios intercostales superiores.
Los pares homolaterales de arterias intercostales anteriores:
- Irrigan las porciones anteriores de los nueve espacios intercostales superiores.
- Discurren lateralmente en el espacio intercostal, una cerca del borde inferior de la costilla superior y la otra cerca del borde superior de la costilla inferior.
- De los dos primeros espacios intercostales se sitúan inicialmente en la fascia endotorácica entre la pleura parietal y los músculos intercostales internos.
- Las que irrigan los espacios intercostales 3º-6º están separadas por tiras del músculo transverso del tórax.
- Las de los espacios intercostales 7º-9º proceden de las arterias musculofrénicas, que también son ramas de las arterias torácicas internas.
- Irrigan los músculos intercostales y dan ramas que, a través de ellos, van a irrigar los músculos pectorales, las mamas y la piel.
- Faltan en los dos espacios intercostales inferiores, que están irrigados únicamente por las arterias intercostales posteriores y sus ramas colaterales.
Venas de la pared torácica
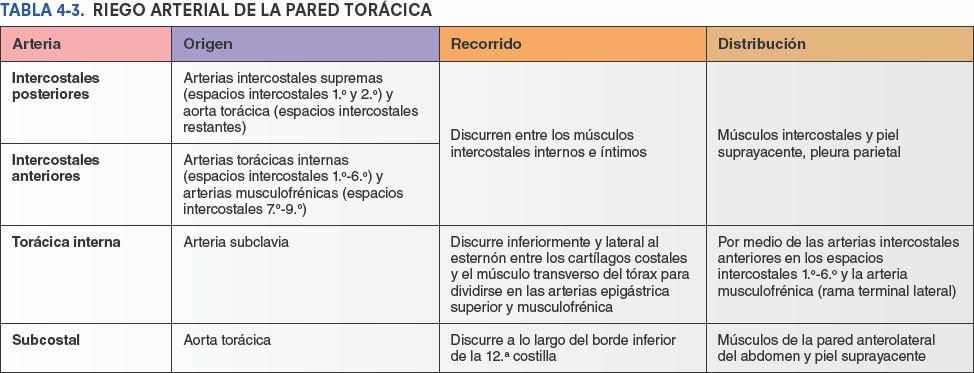
Las venas intercostales acompañan a las arterias y a los nervios intercostales y se sitúan más superiores en los surcos de las costillas (figs. 4-15 B y 4-20). A cada lado hay 11 venas intercostales posteriores y una vena subcostal. Las venas intercostales posteriores se anastomosan con las venas intercostales anteriores (tributarias de las venas torácicas internas). A medida que se aproximan a la columna vertebral, las venas intercostales posteriores reciben una rama posterior, que acompaña al ramo posterior del nervio espinal de ese nivel, y una vena intervertebral que drena los plexos venosos vertebrales asociados a la columna vertebral. Las mayoría de las venas intercostales posteriores (4-11) termina en el sistema venoso ácigos/hemiácigos, que conduce sangre venosa hacia la vena cava superior (VCS).
Las venas intercostales posteriores del primer espacio intercostal suelen entrar directamente en las venas braquiocefálicas derecha e izquierda. Las venas intercostales posteriores de los espacios intercostales 2º y 3º (y algunas veces el 4º) se unen formando un tronco, la vena intercostal superior (fig. 4-20).
La vena intercostal superior derecha es habitualmente la tributaria final de la vena ácigos, antes de que desemboque en la VCS. La vena intercostal superior izquierda, sin embargo, drena normalmente en la vena braquiocefálica izquierda. Esto obliga a la vena a pasar anteriormente a lo largo del lado izquierdo del mediastino superior, en concreto cruzando el arco de la aorta o la raíz de los grandes vasos que nacen de ella, y entre los nervios vago y frénico (fig. 4-71 B). A menudo recibe las venas bronquiales izquierdas y puede recibir también la vena pericardiofrénica izquierda. Típicamente, comunica inferiormente con la vena hemiácigos accesoria. Las venas torácicas internas son las venas satélites de las arterias torácicas internas.
Mamas
Las mamas son los elementos superficiales más destacados de la pared anterior del tórax, especialmente en la mujer. Se componen de tejido glandular y de tejido fibroso de soporte integrados en una matriz de tejido graso, junto con vasos sanguíneos y linfáticos, y nervios. Tanto hombres como mujeres tienen mamas; generalmente sólo están bien desarrolladas en las mujeres (figs. 4-21 y 4-22). Las glándulas mamarias están situadas en el tejido subcutáneo que recubre los músculos pectorales mayor y menor. El pezón se encuentra en la prominencia más elevada de la mama, rodeado por un área circular de piel pigmentada, la areola.
En las mujeres, las glándulas mamarias de las mamas son estructuras accesorias para la reproducción. En el hombre son rudimentarias y carecen de función; consisten sólo en unos pocos conductos pequeños o cordones epiteliales. Normalmente, la grasa de las mamas masculinas no es diferente de la del tejido subcutáneo de cualquier otra localización, y por lo general el sistema glandular no se desarrolla.
Mamas femeninas
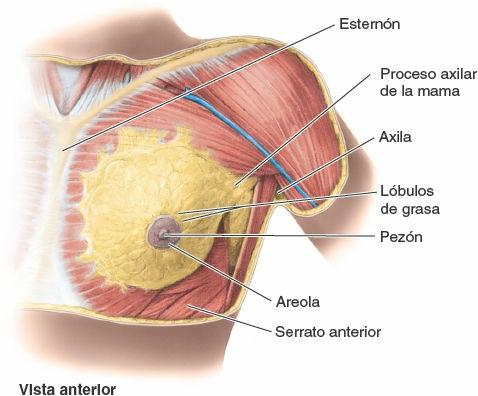
El tamaño de las mamas de una mujer que no amamanta depende de la cantidad de grasa que rodea el tejido glandular. El cuerpo más o menos circular de las mamas femeninas descansa en el lecho de la mama, que se extiende transversalmente desde el borde lateral del esternón hacia la línea axilar media, y verticalmente desde la 2ª hasta la 6ª costillas. Dos terceras partes del lecho de la mama están formadas por la fascia pectoral que cubre el pectoral mayor, y el tercio restante por la fascia que cubre el serrato anterior. Entre la mama y la fascia pectoral se sitúa un plano de tejido subcutáneo laxo o espacio potencial —el espacio (bolsa) retromamario—. Este plano contiene una pequeña cantidad de grasa, y permite a la mama cierto grado de movimiento sobre la fascia pectoral. Una pequeña porción de la glándula mamaria puede extenderse a lo largo del borde inferolateral del pectoral mayor hacia la fosa axilar (axila) y formar el proceso axilar o cola (cola o proceso de Spence). El proceso axilar puede agrandarse durante el ciclo menstrual.
Las glándulas mamarias están unidas con firmeza a la dermis de la piel que las recubre, en especial por ligamentos cutáneos consistentes, los ligamentos suspensorios (de Cooper). Estas condensaciones de tejido conectivo fibroso, particularmente bien desarrolladas en la porción superior de la glándula, ayudan a sostener los lóbulos y lobulillos de la glándula mamaria.
Durante la pubertad (8-15 años de edad), normalmente las mamas crecen, debido en parte al desarrollo glandular pero sobre todo por el aumento del depósito de grasa. Las areolas y los pezones también crecen. El tamaño de la mama y su forma vienen determinados parcialmente por factores genéticos, raciales y dietéticos. Los conductos lactíferos dan lugar a yemas que se desarrollan en 15-20 lóbulos de la glándula mamaria, que constituyen el parénquima (tejido funcional) de la glándula mamaria. Por tanto, cada lóbulo está drenado por un conducto lactífero, y todos ellos convergen para abrirse independientemente. Cada conducto tiene una porción dilatada profunda a la areola, el seno lactífero, donde se acumulan gotitas de leche en la madre lactante. Cuando el recién nacido empieza a mamar, la compresión de la areola (y de los senos lactíferos que hay debajo) exprime las gotitas acumuladas y le estimula a continuar mamando mientras se establece el reflejo de bajada de la leche mediado hormonalmente. La leche de la madre es secretada en la boca del lactante, y no aspirada de la glándula.
Las areolas contienen abundantes glándulas sebáceas, que se dilatan durante el embarazo y secretan una sustancia oleosa que proporciona un lubricante protector para la areola y el pezón. Estas estructuras están especialmente expuestas al roce y la irritación en el inicio de la lactancia materna. Los pezones (papilas mamarias) son prominencias de forma cónica o cilíndrica situadas en el centro de la areola. Los pezones no tienen grasa, pelo ni glándulas sudoríparas. Las puntas de los pezones están fisuradas por los conductos lactíferos que desembocan en ellos. Los pezones están compuestos sobre todo por fibras musculares lisas dispuestas de forma circular que comprimen los conductos lactíferos durante la lactancia y producen la erección de los pezones como respuesta a estímulos, como cuando el lactante empieza a succionar.
Las glándulas mamarias son glándulas sudoríparas modificadas; por consiguiente, no tienen ni cápsula ni vaina. El contorno redondeado y la mayor parte del volumen de las mamas se deben a la grasa subcutánea, excepto durante el embarazo, cuando crecen las glándulas mamarias y se forma nuevo tejido glandular. Los alvéolos (del latín, pequeños espacios huecos) secretores de leche están dispuestos en racimo. En algunas mujeres, las mamas pueden aumentarse de tamaño y volverse dolorosas durante la fase tardía (lútea) del ciclo menstrual. Estos cambios son más probables durante la proliferación de los tejidos glandulares de la mama causados por los niveles cambiantes de estrógenos y progesterona.
Vascularización de la mama
La irrigación arterial de la mama (fig. 4-23 A y B) deriva de:
- Ramas mamarias mediales de las ramas perforantes y ramas intercostales anteriores de la arteria torácica interna, que se origina en la arteria subclavia.
- Arterias torácica lateral y toracoacromial, ramas de la arteria axilar.
- Arterias intercostales posteriores, ramas de la aorta torácica en los espacios intercostales 2º, 3º y 4º.
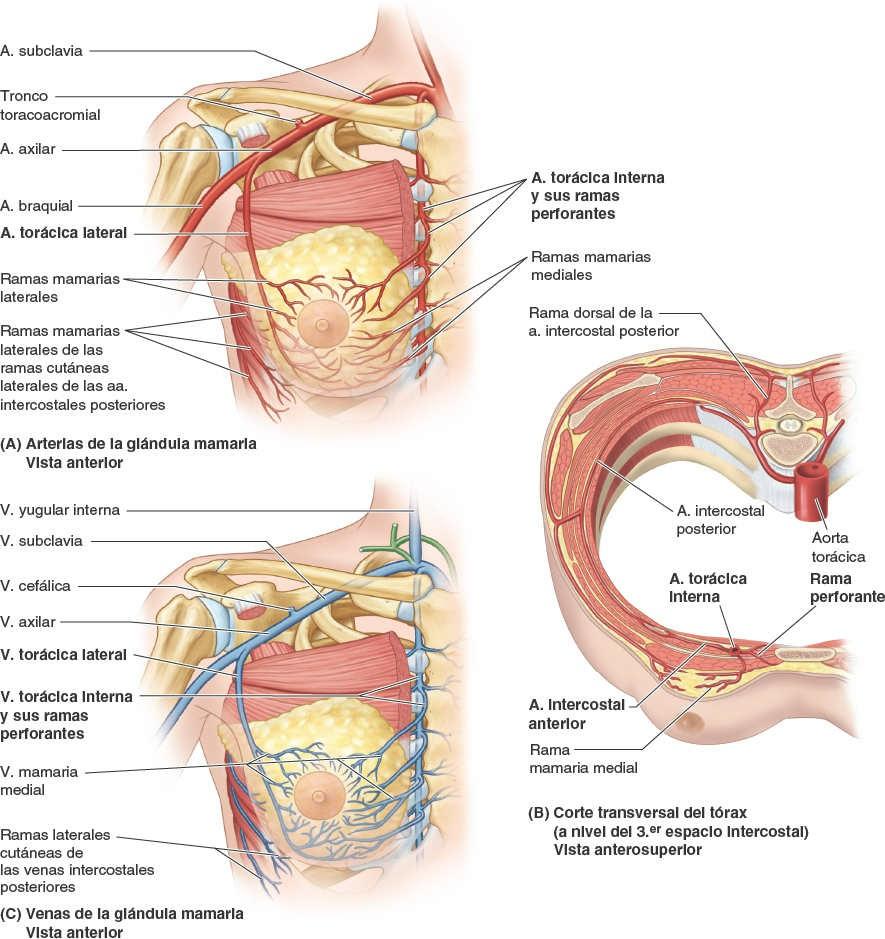
El drenaje venoso de la mama se dirige principalmente a la vena axilar, aunque también drena una parte en la vena torácica interna (fig. 4-23 C).
El drenaje linfático de la mama es importante debido a su papel en las metástasis de células cancerosas. La linfa pasa desde el pezón, la areola y los lóbulos de la glándula hacia el plexo linfático subareolar (fig. 4-24 A y B), y desde ahí:
- La mayor parte de la linfa (> 75%), en especial la de los cuadrantes laterales de las mamas, drena hacia los nódulos linfáticos axilares, inicialmente en su mayoría a los nódulos pectorales o anteriores. No obstante, una porción de linfa puede drenar directamente a otros nódulos axilares, o incluso a los nódulos interpectorales, deltopectorales, supraclaviculares o cervicales profundos inferiores.
- La mayor parte de la linfa restante, en particular la de los cuadrantes mamarios mediales, drena hacia los nódulos linfáticos paraesternales o hacia la mama opuesta, mientras que la linfa de los cuadrantes inferiores puede pasar profundamente hacia nódulos linfáticos abdominales (nódulos linfáticos frénicos inferiores subdiafragmáticos).
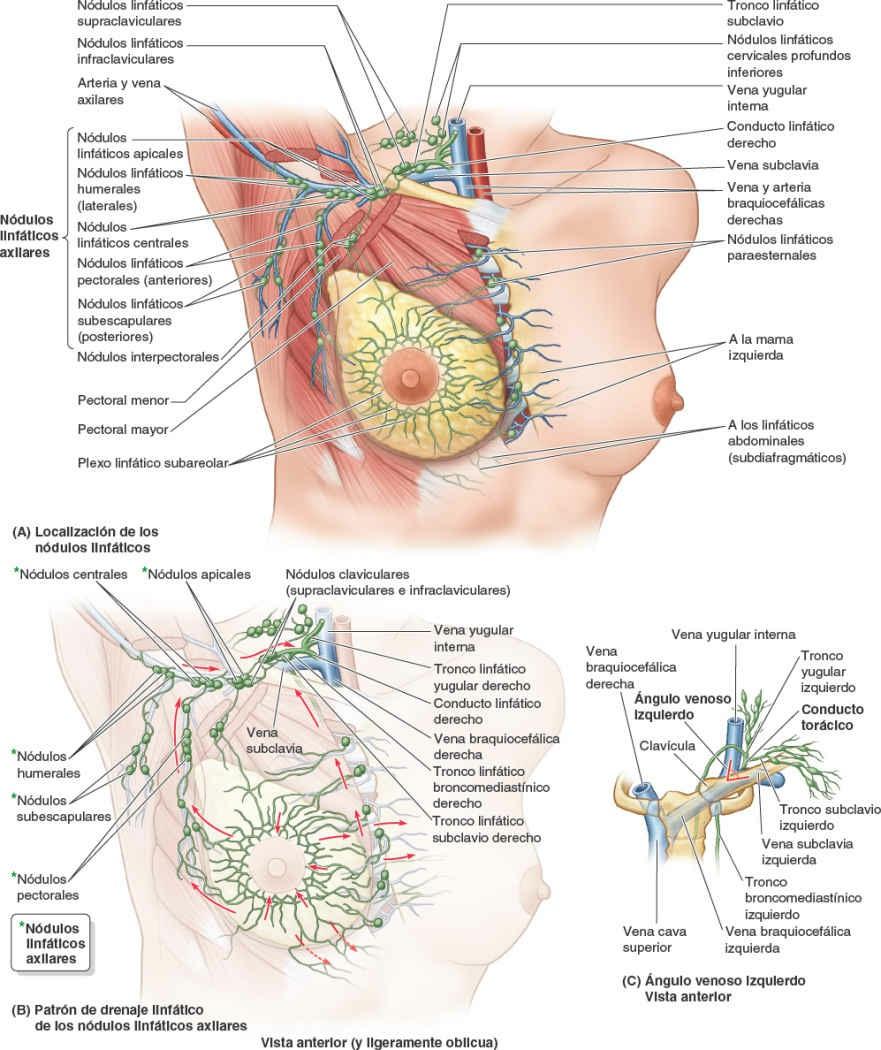
La linfa de la piel de la mama, excepto la de la areola y el pezón, drena en los nódulos linfáticos axilares, cervicales profundos inferiores e infraclaviculares homolaterales, y en los nódulos linfáticos paraesternales de ambos lados.
La linfa de los nódulos axilares drena en los nódulos linfáticos claviculares (supraclaviculares e infraclaviculares) y desde estos en el tronco linfático subclavio, que también drena la linfa del miembro superior. La linfa de los nódulos paraesternales entra en los troncos linfáticos broncomediastínicos, que drenan también la linfa de las vísceras torácicas. La terminación de estos troncos linfáticos es variable; tradicionalmente se ha descrito la fusión de estos troncos entre sí y con el tronco linfático yugular, drenando la cabeza y el cuello para formar un corto conducto linfático derecho en el lado derecho o entrando su terminación en el conducto torácico en el lado izquierdo. Sin embargo, en muchos casos (si no en la mayoría) los troncos desembocan independientemente en la unión de las venas yugular interna y subclavia, los ángulos venosos derecho o izquierdo, que forman las venas braquiocefálicas derecha e izquierda (fig. 4-24 C). En algunas ocasiones, desembocan en ambas venas contribuyentes justo antes del ángulo.
Nervios de la mama
Los nervios de la mama derivan de ramos cutáneos anteriores y laterales de los nervios intercostales 4º-6º (fig. 4-15). Estos ramos de los nervios intercostales atraviesan la fascia pectoral que recubre el pectoral mayor para alcanzar el tejido subcutáneo y la piel de la mama. Los ramos de los nervios intercostales conducen fibras sensitivas de la piel de la mama y fibras simpáticas hasta los vasos sanguíneos de las mamas y el músculo liso en la piel que las recubre y los pezones.
Anatomía de superficie de la pared torácica
Las clavículas se sitúan subcutáneamente formando relieves óseos en la unión del tórax y el cuello (fig. 4-25 A y B). Pueden palparse fácilmente en toda su longitud, sobre todo donde sus extremidades mediales se articulan con el manubrio del esternón. Las clavículas delimitan la división superior entre zonas de drenaje linfático: por encima de las clavículas la linfa fluye en última instancia hacia los nódulos inferiores (nódulos cervicales laterales profundos inferiores) linfáticos yugulares; por debajo de ellas, la linfa parietal (la que procede de la pared del tronco y de los miembros superiores) fluye hacia los nódulos linfáticos axilares.
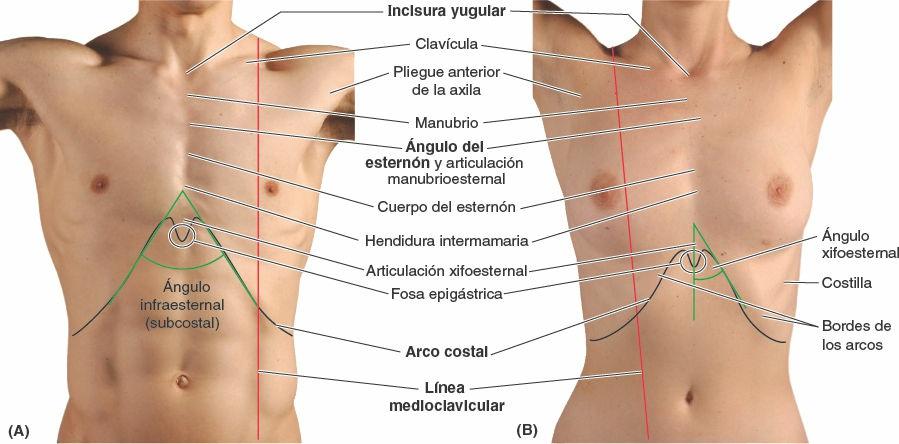
El esternón se sitúa a nivel subcutáneo en la línea media anterior y es palpable a lo largo de toda su longitud. Entre las prominencias de los extremos mediales de las clavículas y las articulaciones esternoclaviculares puede palparse la incisura yugular en el manubrio. La incisura se sitúa al nivel del borde inferior del cuerpo de la vértebra T2 y el espacio entre el primer y el 2º procesos espinosos torácicos.
El manubrio, de unos 4 cm de longitud, se sitúa a nivel de los cuerpos de las vértebras T3 y T4 (fig. 4-26). El ángulo del esternón es palpable, y a menudo visible en las personas jóvenes, debido al ligero movimiento que se produce en la articulación manubrioesternal durante la respiración forzada. El ángulo del esternón se sitúa al nivel del disco intervertebral T4-T5 y el espacio entre el tercer y el 4º procesos espinosos torácicos. El ángulo del esternón señala el nivel del 2º par de cartílagos costales. El lado izquierdo del manubrio es anterior al arco de la aorta, y su lado derecho se sitúa cubriendo directamente la unión de las venas braquiocefálicas para formar la vena cava superior (VCS) (fig. 4-24 C). Debido a que la colocación de catéteres en la VCS para nutrición endovenosa es una práctica clínica frecuente en enfermedades muy graves y para otros propósitos, es fundamental conocer la anatomía de superficie de esta gran vena. La VCS pasa inferiormente profunda al manubrio del esternón y la unión manubrioesternal, pero se proyecta como máximo la anchura de un dedo a la derecha del borde del manubrio. La VCS entra en el atrio (aurícula) derecho del corazón frente al tercer cartílago costal derecho.
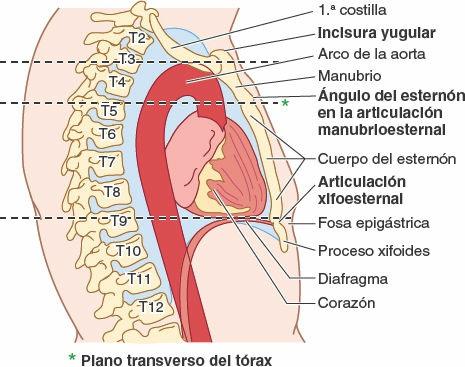
El cuerpo del esternón, de unos 10 cm de longitud, se sitúa anterior al borde derecho del corazón y las vértebras T5-T9 (fig. 4-26). El surco intermamario (depresión en la línea media o separación entre las mamas de las mujeres) se sitúa sobre el cuerpo esternal (figs. 4-25 y 4-29). El proceso xifoides se sitúa en una leve depresión, la fosa epigástrica. Esta fosa se utiliza como guía en la reanimación cardiopulmonar (RCP) para colocar correctamente la mano sobre la parte inferior del esternón. La articulación xifoesternal es palpable y a menudo puede observarse como una cresta, al nivel del borde inferior de la vértebra T9.
Los arcos costales, formados por la unión de los cartílagos costales de las costillas 7ª-10ª, se pueden palpar con facilidad en el punto en que se extienden inferolateralmente desde la articulación xifoesternal. Los cartílagos costales derecho e izquierdo convergen para formar el ángulo infraesternal.
Las costillas y los espacios intercostales proporcionan la base para localizar o describir la posición de estructuras, traumatismos o afectación de la pared torácica o en su profundidad. Debido a que la 1ª costilla no es palpable, el recuento de costillas en las exploraciones físicas comienza en la 2ª, adyacente al ángulo del esternón, que es subcutáneo y se palpa fácilmente. Para contar las costillas y los espacios intercostales anteriormente, se deslizan los dedos lateralmente desde el ángulo del esternón hacia el 2º cartílago costal, y se empiezan a contar las costillas y los espacios moviendo los dedos a partir de ahí. El primer espacio intercostal es el superior al 2º cartílago costal —esto es, los espacios intercostales se numeran de acuerdo con la costilla que forma su límite superior. En general, es más fiable contar los espacios intercostales, ya que la yema del dedo tiende a descansar en (deslizarse hacia) los huecos entre las costillas. Hay que dejar un dedo en ese espacio mientras se utiliza el otro para localizar el espacio siguiente. Si utilizamos todos los dedos es posible localizar cuatro espacios al mismo tiempo. Los espacios son más anchos anterolateralmente (aproximadamente en la línea medioclavicular). Si se separan los dedos de la pared torácica mientras se cuentan los espacios, es fácil que el dedo vuelva a un mismo espacio y se confunda con el de abajo. Posteriormente, el extremo medial de la espina de la escápula se sitúa sobre la 4ª costilla.
Mientras que las costillas y/o los espacios intercostales proporcionan la «latitud» para la orientación y la localización en la pared torácica, diversas líneas imaginarias facilitan las descripciones anatómicas y clínicas al proporcionar la «longitud». Las líneas mencionadas a continuación están extrapoladas sobre la pared torácica basándose en características superficiales visibles o palpables:
- La línea media anterior (medioesternal) indica la intersección del plano medio con la pared anterior del tórax (fig. 4-27 A).
- La línea medioclavicular pasa a través del punto medio de la clavícula, paralela a la línea media anterior.
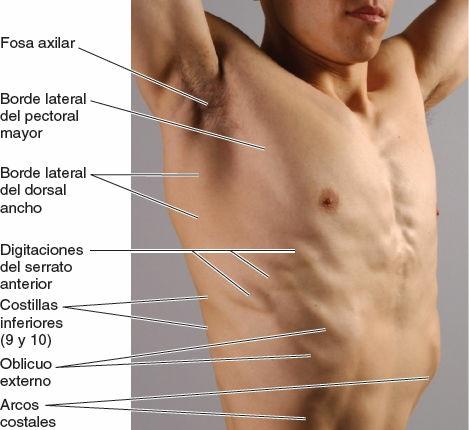
- La línea axilar anterior discurre verticalmente a lo largo del pliegue anterior de la axila, que está formado por el borde inferolateral del músculo pectoral mayor a medida que se expande desde el tórax hasta el húmero en el brazo (fig. 4-27 B).
- La línea axilar media discurre desde el vértice (parte más profunda) de la axila, paralela a la línea axilar anterior.
- La línea axilar posterior, también paralela a la línea axilar anterior, discurre verticalmente a lo largo del pliegue posterior de la axila formado por los músculos dorsal ancho y redondo mayor, a medida que se expanden desde el dorso hacia el húmero.
- La línea media posterior (mediovertebral) es una línea vertical que discurre a lo largo de los vértices de los procesos espinosos de las vértebras (fig. 4-27 C).
- Las líneas escapulares son paralelas a la línea media posterior y cruzan los ángulos inferiores de las escápulas.
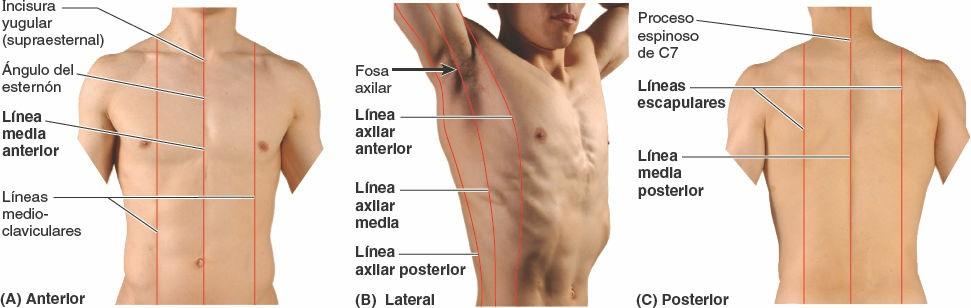
Se extrapolan otras líneas (que no se muestran) a lo largo de los bordes palpables de formaciones óseas como las líneas paraesternal y paravertebral a cada lado del esternón y la columna vertebral.
Las mamas son los elementos superficiales más destacados de la pared anterior del tórax, especialmente en la mujer. Excepto en caso de un exceso de tejido subcutáneo, en el hombre las mamas son en general una acentuación del contorno de los músculos pectorales mayores, realzado por la presencia del pezón en el 4º espacio intercostal, lateral a la línea medioclavicular (fig. 4-28). En los individuos moderadamente atléticos, el contorno de los músculos pectorales mayores está bien definido, y están separados en la línea media por el surco intermamario sobre el esternón, con el borde lateral formando el pliegue axilar anterior (fig. 4-25). Inferolateralmente, unas bandas similares a un dedo, o digitaciones del serrato mayor, tienen un aspecto de dientes de sierra insertándose en las costillas e interdigitándose con el oblicuo externo del abdomen (fig. 4-28). Las costillas inferiores y los arcos costales a menudo son evidentes, en especial cuando los músculos abdominales están contraídos. La musculatura intercostal no suele observarse normalmente; no obstante, en algún caso (raramente) en que hay ausencia o atrofia de la musculatura intercostal, los espacios intercostales se hacen evidentes con la respiración: durante la inspiración, son cóncavos y durante la espiración protruyen.
Las mamas femeninas tienen un tamaño, una forma y una simetría variables — incluso en una misma mujer. Sus superficies anteriores aplanadas no muestran una demarcación clara con la superficie anterior de la pared torácica, pero sus bordes están bien definidos lateralmente e inferiormente (fig. 4-29). A menudo es visible un patrón venoso sobre las mamas, sobre todo durante el embarazo.
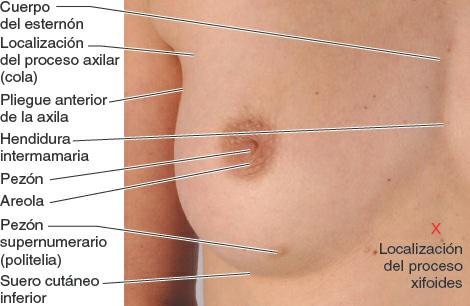
El pezón está rodeado por un área pigmentada, ligeramente sobreelevada y circular, la areola, cuyo color varía con la complexión de la mujer. La areola generalmente se oscurece durante el embarazo y a partir de entonces mantiene ese color. La areola suele estar punteada por aberturas papulares (levemente elevadas) de las glándulas areolares (glándulas sebáceas de la piel de la areola). En ocasiones, uno o los dos pezones están invertidos (retraídos); esta leve anomalía congénita puede dificultar la lactancia.
En los hombres y en las mujeres nulíparas jóvenes —las que nunca han dado a luz un feto viable— con mamas de un tamaño moderado, el pezón se sitúa anterior al 4º espacio intercostal, a unos 10 cm de la línea media anterior. En general, los pezones muestran una considerable variabilidad con el tamaño de la mama, sobre todo en las mujeres multíparas. Por consiguiente, y debido a las variaciones en el tamaño y en la forma, los pezones no son una referencia fiable del 4º espacio intercostal en las mujeres adultas.