03. Función del corazón como bomba
El corazón es una bomba muscular de 4 cámaras con tamaño aproximado al del puño de un hombre. Late en promedio 70 veces por minuto las 24 h del día, los 365 días de cada año durante toda la vida. En un día, esta bomba desplaza más de 6.800 l de sangre por todo el cuerpo.
Anatomía funcional del corazón
El corazón se localiza entre los pulmones, en el espacio mediastínico de la cavidad torácica dentro de un saco laxo llamado pericardio. Está suspendido por los grandes vasos, con el lado más ancho (es decir, la base) hacia arriba y el vértice es decir, dirigido hacia abajo, al frente y a la izquierda.
El corazón está en posición oblicua, de manera que el lado derecho está casi frente al lado izquierdo del corazón y sólo una pequeña porción de la cara lateral del ventrículo izquierdo está en el plano frontal del corazón (figura 29-3). Cuando se coloca la mano sobre el tórax, el impacto principal de la contracción cardíaca se percibe contra la pared torácica en un punto entre la quinta y sexta costillas, un poco debajo del pezón y unos 7,5 cm a la izquierda de la línea media. Este se llama punto de impulso máximo (PIM).
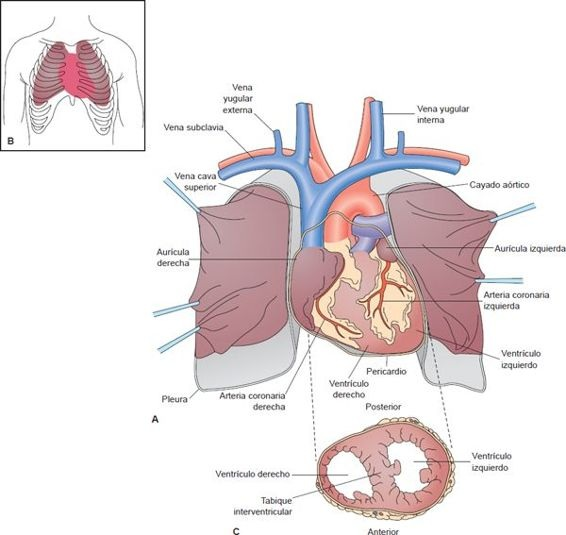
La pared del corazón está compuesta por el epicardio externo, que recubre la cavidad pericárdica; el miocardio o capa muscular; y el endocardio suave, que recubre las cámaras cardíacas (figura 29-4). Un esqueleto fibroso sostiene las estructuras vasculares del corazón. Los tabiques interauricular e interventricular dividen al corazón en una bomba derecha y una izquierda, cada una formada por 2 cámaras musculares: una aurícula de paredes delgadas que sirve como reservorio para la sangre que llega al corazón y un ventrículo de paredes gruesas que bombea la sangre fuera del corazón. El mayor grosor de la pared ventricular izquierda se debe al trabajo adicional que debe realizar este ventrículo para desplazar la sangre oxigenada a las extremidades.
Pericardio
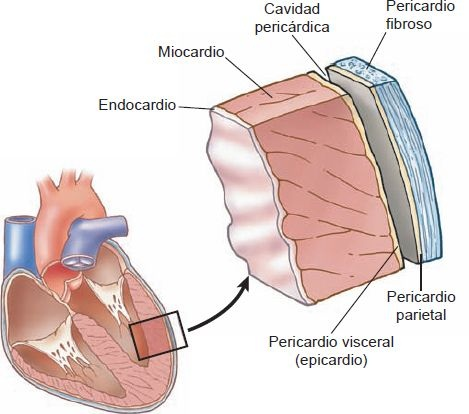
El pericardio forma una cubierta fibrosa alrededor del corazón, lo mantiene en una posición fija en el tórax y brinda protección física, además de ser una barrera a la infección. El pericardio consiste en una fuerte capa fibrosa externa y una delgada capa serosa interna. La capa fibrosa externa está unida con los grandes vasos que entran y salen del corazón, con el esternón y el diafragma. El pericardio fibroso es muy resistente a la distensión; impide la dilatación aguda de las cámaras cardíacas y ejerce un efecto restrictivo sobre el ventrículo izquierdo. La capa serosa interna consiste en un manto visceral y uno parietal.
La capa visceral, también conocida como epicardio, cubre todo el corazón y los grandes vasos, luego se pliega para formar la capa parietal que recubre el pericardio fibroso (figura 29-4). Entre las capas visceral y parietal está la cavidad pericárdica, un espacio potencial que contiene 30 ml a 50 ml de líquido seroso. Este líquido actúa como lubricante para minimizar la fricción con las estructuras circundantes cuando el corazón se contrae y se relaja.
Miocardio
El miocardio, o porción muscular del corazón, forma las paredes de las aurículas y los ventrículos. Al igual que el músculo esquelético, las células musculares cardíacas son estriadas y están formadas por sarcómeras que contienen filamentos de actina y miosina. Son más pequeñas y compactas que las células del músculo esquelético y contienen muchas mitocondrias grandes, reflejo de sus necesidades continuas de energía.
Las propiedades contráctiles del músculo cardíaco son similares a las del músculo esquelético, excepto que las contracciones son involuntarias y la duración de la contracción es mucho más prolongada. A diferencia de la disposición ordenada longitudinal de las fibras musculares esqueléticas, las células del músculo cardíaco están dispuestas como un enrejado interconectado en el que las fibras se dividen, se recombinan y luego se dividen de nuevo (figura 29-5A). Las fibras están separadas de las células musculares vecinas por estructuras densas llamadas discos intercalados. Estos discos, únicos del músculo cardíaco, contienen uniones comunicantes que sirven como vías de baja resistencia para el paso de iones e impulsos eléctricos de una célula cardíaca a otra (figura 29-5B).
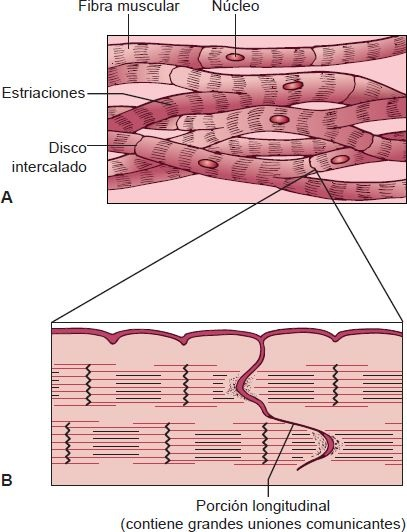
Por tanto, el miocardio se comporta como una unidad, o sincitio, en lugar de un grupo de unidades aisladas, como el músculo esquelético. Cuando se estimula una célula miocárdica, el impulso viaja con rapidez, de manera que el corazón puede latir como unidad. El músculo cardíaco tiene 2 sincitios: el auricular y el ventricular. El sincitio auricular constituye las paredes de las aurículas y el sincitio ventricular, las paredes de los ventrículos. Estos 2 tipos de sincitio permiten que las aurículas se contraigan antes que los ventrículos, lo cual es importante para el bombeo cardíaco.
Como en el músculo esquelético, la contracción miocárdica se basa en los filamentos de actina y miosina, que interactúan y se deslizan entre sí durante la contracción. Varias proteínas importantes regulan la unión entre actina y miosina, e incluyen la tropomiosina y el complejo de la troponina. El complejo de troponina tiene 3 subunidades (troponina T, troponina I y troponina C) que regulan la contracción mediada por calcio en el músculo estriado. En la práctica clínica, la medición de laconcentración sérica de las formas cardíacas de troponina T y troponina I se utiliza en el diagnóstico del infarto de miocardio.
Aunque las células del músculo cardíaco requieren calcio para contraerse, tienen un retículo sarcoplásmico menos definido para almacenar calcio que las células del músculo esquelético. Por tanto, el miocardio depende más de la entrada de iones calcio extracelular para contraerse que el músculo esquelético. Los glucósidos cardíacos (ej. digoxina) son fármacos inotrópicos que aumentan la contractilidad cardíaca mediante el aumento de la concentración de calcio intracelular libre en la proximidad de los filamentos de actina y miosina.
Endocardio
El endocardio es una membrana delgada de 3 capas que recubre el corazón y las válvulas. La capa más interna consiste en células endoteliales lisas sostenidas por una capa delgada de tejido conectivo.
El recubrimiento endotelial del endocardio se continúa con el recubrimiento de los vasos sanguíneos que entran y salen del corazón. La capa intermedia consiste en tejido conectivo denso con fibras elásticas. La capa externa, que está formada por células de tejido conectivo dispuestas en forma irregular, contiene vasos sanguíneos y ramificaciones del sistema de conducción, y se continúa con el miocardio.
Válvulas cardíacas y esqueleto fibroso
Una característica estructural importante del corazón es su esqueleto fibroso, que consiste en 4 anillos valvulares interconectados y el tejido conectivo que los rodea. Separa las aurículas de los ventrículos y forma un soporte rígido para la unión de las válvulas y la inserción del músculo cardíaco (figura 29-6). Las partes superiores de los anillos valvulares se unen con el tejido muscular de las aurículas, troncos pulmonares y aorta. Las partes inferiores se unen con las paredes ventriculares. Para que el funcionamiento del corazón sea efectivo, la sangre debe fluir sólo en una dirección, con desplazamiento anterógrado por las cámaras derechas a los pulmones y luego por las cámaras izquierda a la circulación sistémica (figura 29-7). Este flujo unidireccional se obtiene con el par de válvulas auriculoventriculares (es decir, tricúspide y mitral) y 2 válvulas semilunares (es decir, pulmonar y aórtica).
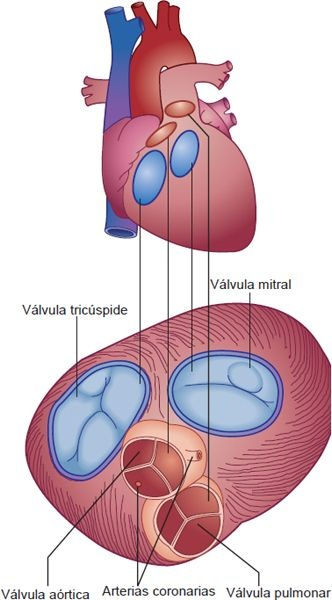
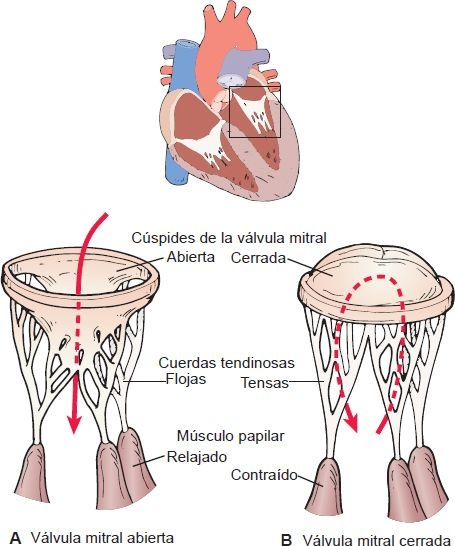
Cuando se cierran, las válvulas AV impiden el reflujo de sangre de los ventrículos a las aurículas durante la sístole. Los bordes delgados de las válvulas AV forman cúspides, 2 del lado izquierdo del corazón (es decir, válvula bicúspide o mitral) y 3 del lado derecho (es decir, válvula tricúspide). La válvula bicúspide también se llama válvula mitral. Las válvulas AV están sostenidas por los músculos papilares, que se proyectan de la pared ventricular, y por las cuerdas tendinosas, que se unen con la válvula (figura 29-8). La contracción de los músculos papilares al inicio de la sístole asegura el cierre al ejercer tensión sobre las valvas de las válvulas AV, antes de la fuerza completa de la contracción ventricular las presione. Las cuerdas tendinosas son estructuras parecidas a cordones que sostienen las válvulas AV e impiden que se eviertan hacia las aurículas durante la sístole.
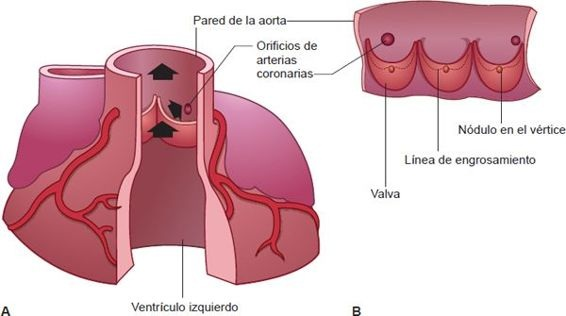
Las válvulas aórtica y pulmonar previenen el reflujo de la aorta y la arteria pulmonar a los ventrículos durante la diástole. La válvula pulmonar, que se localiza entre el ventrículo derecho y la arteria pulmonar, controla el flujo sanguíneo hacia la circulación pulmonar, y la válvula aórtica, situada entra el ventrículo izquierdo y la aorta, controla el flujo de sangre hacia la circulación sistémica. Como sus valvas tienen forma de media luna, a menudo se conocen como válvulas semilunares. Estas válvulas tienen 3 cúspides unidas a los anillos valvulares (figura 29-9B). Estas estructuras semejantes a copas reciben el flujo retrógrado de la sangre que ocurre al final de la sístole, lo que favorece el cierre. Para que se forme un sello perfecto en los 3 bordes de las válvulas semilunares, cada cúspide valvular debe tener una forma triangular, lo que se facilita con un engrosamiento nodular en el vértice de cada valva (figura 29-9A).
Detrás de las válvulas semilunares están los senos de Valsalva. En estos espacios se forman corrientes de remolino que tienden a mantener las cúspides valvulares alejadas de las paredes del vaso. Las aberturas de las arterias coronarias se localizan detrás de las cúspides derecha e izquierda de la válvula aórtica. De no ser por la presencia de los senos de Valsalva y las corrientes de remolino, las aberturas de las arterias coronarias se bloquearían por las cúspides valvulares.
No existen válvulas en los sitios de las aurículas (es decir, venas cavas y venas pulmonares) por donde entra la sangre al corazón. Esto significa que el exceso de sangre se empuja de regreso a las venas cuando las arterias se distienden. Por ejemplo, las venas yugulares casi siempre se vuelven prominentes en la insuficiencia cardíaca derecha grave, pero en condiciones normales se encuentranplanas o colapsadas. De igual manera, el sistema venoso pulmonar se congestiona cuando se impide el flujo de salida de la aurícula izquierda.
Ciclo cardíaco
El término ciclo cardíaco se emplea para describir la acción rítmica de bombeo del corazón. El ciclo cardíaco se divide en 2 partes:
- Sístole, el período en el que los ventrículos se contraen.
- Diástole, el período en el que los ventrículos se relajan y se llenan con sangre.
Durante el ciclo cardíaco, existen cambios simultáneos en la presión auricular izquierda, presión ventricular izquierda, volumen ventricular, ECG y los ruidos cardíacos (figura 29-19). La actividad eléctrica, registrada en la ECG, precede a los sucesos mecánicos del ciclo cardíaco.
La pequeña onda P redondeada de la ECG representa la despolarización del nodo senoauricular (esdecir, el marcapaso del corazón), el tejido de conducción auricular y la masa muscular auricular. El complejo QRS registra la despolarización del sistema de conducción ventricular y la masa muscular ventricular. La onda T de la ECG se produce durante la última mitad de la sístole y representa la repolarización de los ventrículos.
Sístole y diástole ventriculares
La sístole ventricular se divide en 2 períodos: el período de contracción isovolumétrica y el de eyección.
El período de contracción isovolumétrica, que comienza con el cierre de las válvulas AV y con la presencia del primer ruido cardíaco, anuncia el inicio de la sístole. Justo después del cierre de las válvulas AV, hay un intervalo adicional de 0,02 s a 0,03 s en el que las válvulas semilunares de salida (pulmonar y aórtica) permanecen cerradas. Durante este período (figura 29-10), las presiones ventriculares se elevan en forma súbita porque tanto las válvulas AV como las semilunares están cerradas y la sangre no sale de los ventrículos. Los ventrículos continúan la contracción hasta que la presión ventricular izquierda es un poco más alta que la presión aórtica y la presión ventricular derecha es mayor que la presión en la arteria pulmonar. En ese momento, las válvulas semilunares se abren, con lo que inicia el período de eyección. Casi el 60% del volumen por latido se eyecta durante el primer cuarto de la sístole, el 40% restante se eyecta durante los siguientes 2 cuartos de la sístole.
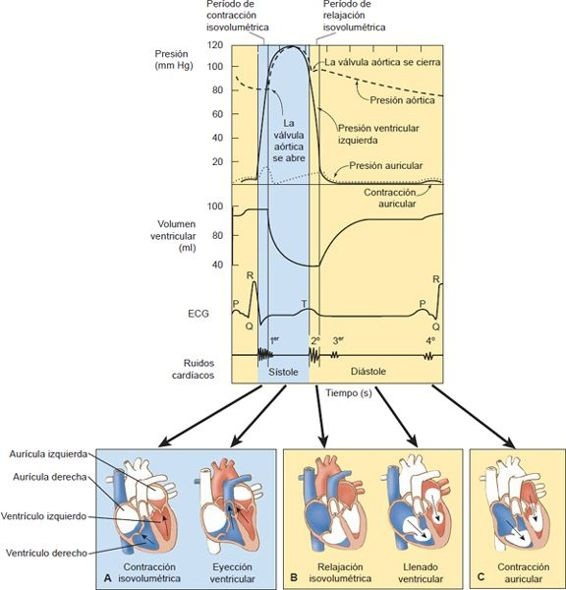
Poca sangre sale del corazón durante el último cuarto de la sístole, aunque los ventrículos permanecen contraídos. Al final de la sístole, los ventrículos se relajan, lo que produce una caída precipitada de la presión intraventricular. Cuando esto ocurre, la sangre de las grandes arterias regresa hacia los ventrículos, lo que hace que las válvulas aórtica y pulmonar se cierren, un fenómeno marcado por el segundo ruido cardíaco.
La presión aórtica refleja cambios en la eyección de sangre por el ventrículo izquierdo. Se produce un aumento en la presión y estiramiento de las fibras elásticas de la aorta cuando la sangre se eyecta a esta arteria al principio de la sístole. La presión aórtica continúa en ascenso y luego empieza a disminuir durante el último cuarto de la sístole, conforme la sangre fluye de la aorta hacia los vasos periféricos. La incisura o muesca en el trazo de la presión aórtica representa el cierre de la válvula aórtica. Esto se debe a un corto período de reflujo sanguíneo justo antes del cierre valvular. La aorta es muy elástica y, por tanto, se estira durante la sístole para recibir la sangre que se eyecta del lado izquierdo del corazón durante la sístole. En la diástole, la recuperación de las fibras elásticas en la aorta sirve para mantener la presión aórtica.
La diástole está marcada por la relajación y llenado de los ventrículos. Después del cierre de las válvulas semilunares, los ventrículos continúan relajados durante 0,03 s a 0,06 s más (denominado período de relajación isovolumétrica). Durante este intervalo, las válvulas semilunares y AV permanecen cerradas, y el volumen ventricular permanece igual, mientras la presión ventricular desciende hasta ser menor que la presión auricular (figura 29-10). Cuando esto ocurre, las válvulas AV se abren y la sangre que se había acumulado en las aurículas durante la sístole fluye hacia los ventrículos. La mayor parte del llenado ventricular ocurre en el primer tercio de la diástole, llamado período de llenado rápido. Durante el tercio medio de la diástole, la entrada a los ventrículos esmínima. El último tercio de la diástole está marcado por la contracción auricular, lo que da un impulso adicional al llenado ventricular y representa cerca del 20% del llenado de los ventrículos.
Cuando es audible, el tercer ruido cardíaco se escucha durante el período de llenado rápido de la diástole, mientras la sangre fluye hacia un ventrículo distendido o no distensible. El cuarto ruido cardíaco se produce en el último tercio de la diástole, cuando las aurículas se contraen.
Durante la diástole, los ventrículos aumentan su volumen hasta cerca de 120 ml (el volumen al final de la diástole). Al final de la sístole, cerca de 40 ml a 50 ml de sangre (el volumen al final de la sístole) permanecen en los ventrículos (figura 29-10). La diferencia entre los volúmenes al final de la diástole y al final de la sístole (cerca de 70 ml) se llama volumen por latido. La fracción de eyección, que es el volumen por latido dividido entre el volumen al final de la diástole, representa la fracción o porcentaje del volumen al final de la diástole que se eyecta del corazón durante la sístole. La fracción de eyección ventricular izquierda (el valor normal del 55% al 75% cuando se determina por ecocardiografía o angiocardiografía) se emplea a menudo para evaluar el pronóstico de las personas con diversas enfermedades cardíacas.
Llenado y contracción auriculares
Existen 3 ondas de presión auricular principales durante el ciclo cardíaco: las ondas a, c y v.
La onda a se produce durante la última parte de la diástole y se produce por la contracción auricular. La onda c ocurre cuando los ventrículos empiezan a contraerse y su presión elevada hace que las válvulas AV se abulten hacia las aurículas. La onda v ocurre hacia el final de la sístole, cuando las válvulas AV todavía están cerradas, se produce por la acumulación lenta de sangre en las aurículas.
Las ondas de presión auricular derecha se transmiten a las venas yugulares internas como pulsaciones. Estas pulsaciones pueden valorarse en forma visual y emplearse para evaluar la función cardíaca. Por ejemplo, las ondas a se exageran cuando aumenta el volumen de la aurícula derecha por alteración del vaciamiento del ventrículo derecho.
Como no hay válvulas en las uniones de las venas centrales (es decir, venas cavas y venas pulmonares) con las aurículas, el llenado auricular ocurre durante la sístole y la diástole. Con la respiración normal y tranquila, la presión auricular derecha casi siempre varía entre −2 y +2 mm Hg. Es esta presión auricular baja la que mantiene el desplazamiento de sangre desde las venas sistémicas a la aurícula derecha y de las venas pulmonares a la aurícula izquierda.
La presión auricular derecha está regulada por el balance entre la capacidad del corazón para desplazar la sangre de las cámaras derechas y por el lado izquierdo del corazón hacia la circulación sistémica, y por la tendencia de la sangre a fluir de la circulación periférica a la aurícula derecha.
Cuando el corazón bombea con fuerza, la presión auricular derecha disminuye y el llenado auricular se incrementa. La presión auricular derecha también se modifica por lo cambios en la presión intratorácica. Disminuye durante la inspiración, cuando la presión intratorácica se vuelve más negativa, y aumenta durante la tos o espiración forzada, cuando la presión intratorácica se hace más positiva. El retorno venoso es un reflejo de la cantidad de sangre en la circulación sistémica que está disponible para el retorno al corazón derecho y de la fuerza que impulsa la sangre de nuevo al lado derecho del corazón. El retorno venoso aumenta cuando el volumen sanguíneo aumenta o cuando la presión auricular cae; se reduce en el shock hipovolémico o cuando la presión auricular se eleva.
Aunque la función principal de las aurículas es almacenar sangre a su entrada al corazón, estas cámaras también actúan como bombas que ayudan al llenado ventricular. La contracción auricular ocurre durante el último tercio de la diástole. La contracción auricular se vuelve más importante durante períodos con aumento de actividad, cuando el tiempo de llenado auricular se reduce por el aumento de la frecuencia cardíaca o cuando alguna enfermedad del corazón afecta el llenado ventricular. En estas 2 situaciones, el gasto cardíaco disminuiría en forma drástica si no fuera por la acción de las aurículas. Se calcula que la contracción auricular puede contribuir hasta con un 20% a la reserva cardíaca en períodos con aumento en la necesidad, pero tiene poco o ningún efecto en el shock cardíaco durante el reposo.
Regulación del funcionamiento cardíaco
La eficiencia y trabajo del corazón como bomba a menudo se mide en términos de gasto cardíaco o la cantidad de sangre que el corazón bombea por minuto. El gasto cardíaco (GC) es el producto del volumen por latido (VL) y la frecuencia cardíaca (FC), y puede expresarse por la ecuación GC = VL × FC. El gasto cardíaco varía según el tamaño del cuerpo y las necesidades metabólicas de los tejidos. Aumenta con la actividad física y disminuye durante el reposo y el sueño. El GC promedio en reposo en los adultos normales varía entre 4 lpm y 6 lpm. En el atleta de élite que realiza un ejercicio extremo, quizá sea necesario que el corazón bombee de 4 a 6 veces esa cantidad.
La reserva cardíaca se refiere al porcentaje máximo de aumento en el gasto cardíaco que puede sobre el nivel normal de reposo. El adulto joven sano tiene una reserva cardíaca aproximada del 300% al 400%. El funcionamiento del corazón está influenciado por las demandas de trabajo y la capacidad de la circulación coronaria para cubrir sus necesidades metabólicas.
La capacidad del corazón para aumentar su gasto de acuerdo con las necesidades del cuerpo depende sobre todo de 4 factores:
- Precarga o llenado ventricular.
- Poscarga o resistencia a la eyección cardíaca de sangre.
- Contractilidad cardíaca.
- Frecuencia cardíaca.
La frecuencia y la contractilidad cardíacas son factores cardíacos estrictos, lo que significa que se originan en el corazón, aunque están controladas por varios mecanismos neurales y humorales. Por otra parte, la precarga y la poscarga dependen mutuamente de la actividad del corazón y la vasculatura. No sólo determinan el gasto cardíaco: ellas mismas están determinadas por el gasto cardíaco y por ciertas características vasculares.
Precarga
La precarga es el trabajo volumétrico del corazón. Casi siempre se considera la presión al final de la diástole cuando el ventrículo está lleno. Se llama precarga porque es el trabajo o carga impuestos al corazón antes del comienzo de la la contracción. Es la cantidad de sangre que el corazón debe bombear con cada latido. Depende sobre todo del retorno venoso al corazón y el estiramiento concurrente de las fibras miocárdicas.
El aumento en la fuerza de contracción que acompaña a un aumento en el volumen ventricular al final de la diástole se denomina mecanismo de Frank-Starling o ley de Starling del corazón (figura 29-11). La disposición anatómica de los filamentos de actina y miosina en las fibras miocárdicas es tal, que la tensión o fuerza de contracción depende del grado de estiramiento de las fibras musculares justo antes que los ventrículos empiecen a contraerse. La fuerza de contracción y el gasto cardíaco máximos se alcanzan cuando el retorno venoso produce un aumento en el llenado ventricular al final de la diástole (es decir, la precarga), de manera que las fibras musculares se estiren 2,5 veces su longitud de reposo normal. Cuando las fibras musculares se estiran en esta medida, se produce la superposición óptima de los filamentos de actina y miosina para la contracción máxima.
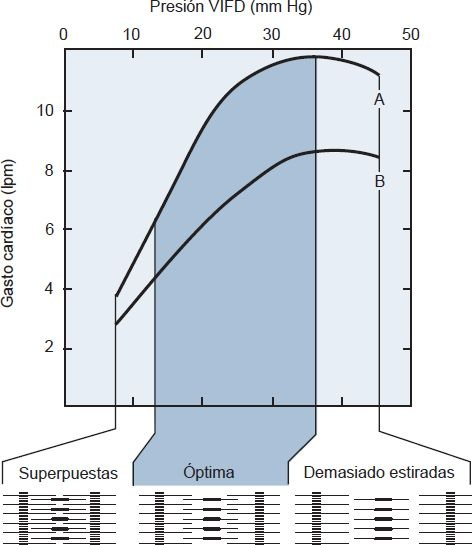
El mecanismo de Frank-Starling permite que el corazón ajuste su capacidad de bombeo para aceptar el retorno venoso de varias magnitudes. Cuando una mayor cantidad de sangre fluye a los ventrículos, el músculo cardíaco se estira más. El gasto cardíaco es menor cuando el llenado insuficiente produce una superposición excesiva de los filamentos de actina y miosina, o cuando el llenado excesivo hace que los filamentos se separen demasiado.
Poscarga
La poscarga es la presión en la que el músculo ejerce su fuerza contráctil para desplazar la sangre hacia la aorta. Se llama poscarga porque es el trabajo que se impone al corazón después del inicio de la contracción. La presión arterial sistémica es la principal fuente de poscarga para el corazón izquierdo y la presión arterial pulmonar lo es para el corazón derecho. El trabajo de poscarga del ventrículo izquierdo también aumenta con el estrechamiento (es decir, estenosis) de la válvula aórtica. Por ejemplo, en las etapas tardías de la estenosis aórtica, es posible que el ventrículo izquierdo necesite generar presiones sistólicas de hasta 300 mm Hg para impulsar la sangre a través de la válvula alterada.
Contractilidad cardíaca
La contractilidad cardíaca se refiere a la capacidad del corazón para cambiar su fuerza de contracción sin modificar su longitud de reposo (es decir, diastólica). El estado contráctil del miocardio depende de las propiedades bioquímicas y biofísicas que regulan las interacciones de la actina y la miosina en las células miocárdicas. Depende mucho de la cantidad de iones calcio que estén disponibles para participar en el proceso contráctil.
Una influencia inotrópica es la que modifica el estado contráctil del miocardio de manera independiente al mecanismo de Frank-Starling (figura 29-11, curva superior). Por ejemplo, la estimulación simpática tiene un efecto inotrópico positivo porque aumenta el calcio disponible para la interacción entre los filamentos de actina y miosina. La hipoxia ejerce un efecto inotrópico negativo porque interfiere con la generación de trifosfato de adenosina (ATP), necesario para la contracción muscular.
Frecuencia cardíaca
La frecuencia cardíaca determina la periodicidad con la que la sangre se eyecta del corazón. Por lo tanto, conforme aumenta la frecuencia cardíaca, el gasto cardíaco tiende a aumentar. Con el aumento de la frecuencia cardíaca se reduce el tiempo disponible para la diástole y los ventrículos tienen menos tiempo para llenarse. A una frecuencia cardíaca de 75 lpm, un ciclo cardíaco dura 0,8 s, de los cuales cerca de 0,3 s se dedican a la sístole y quedan alrededor de 0,5 s para la diástole. Conforme la frecuencia cardíaca aumenta, el tiempo de la sístole permanece casi igual y el de la diástole se reduce. Esto disminuye el volumen por latido y, con frecuencias cardíacas elevadas, el gasto cardíaco es menor. Uno de los peligros de la taquicardia ventricular es la reducción del gasto cardíaco porque el corazón no tiene tiempo para llenarse de manera adecuada.