01. Organización estructural del sistema respiratorio
El sistema respiratorio consiste en los pasajes de aire (2 pulmones) y los vasos sanguíneos que los alimentan. Consta también de las estructuras que proporcionan un mecanismo ventilador, es decir, la caja torácica y los músculos respiratorios, que incluyen el diafragma y el músculo respiratorio principal.
Los pulmones son órganos blandos, esponjosos, en forma de cono, localizados lado a lado en la cavidad torácica (figura 35-1). Están separados entre sí por el mediastino (es decir, el espacio entre los pulmones) y su contenido: el corazón, vasos sanguíneos, nodos linfáticos, fibras nerviosas, glándula del timo y esófago.
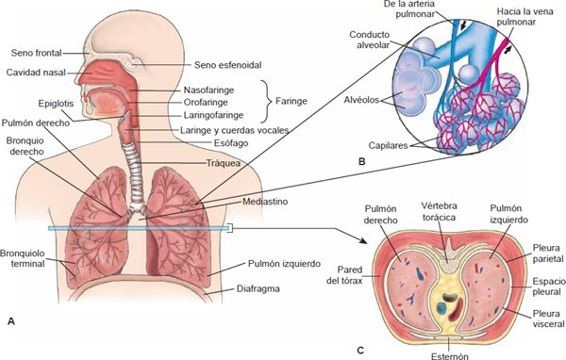
La parte superior del pulmón, que descansa en la parte superior de la cavidad torácica, se llama ápice, y la parte inferior, que se apoya en el diafragma, se conoce como la base. Los pulmones se dividen en lóbulos, 3 en el pulmón derecho y 2 en el izquierdo.
Funcionalmente, el sistema respiratorio puede dividirse en 2 partes: las vías respiratorias conductoras, por las que se mueve el aire a medida que pasa entre la atmósfera y los pulmones, y los tejidos respiratorios de los pulmones, en donde tiene lugar el intercambio de gas.
Vías respiratorias de conducción
Las vías respiratorias de conducción consisten en los pasajes nasales, boca y faringe, laringe, tráquea, bronquios y bronquiolos (figura 35-1). Además de funcionar como un conducto para flujo de aire, las vías respiratorias de conducción sirven para «acondicionar» el aire inspirado. El aire que respiramos se calienta, filtra y humedece a medida que se mueve por estas estructuras. El calor se transfiere al aire desde la sangre que fluye por las paredes de los pasajes respiratorios. La capa mucociliar remueve materia extraña y el agua de las membranas mucosas se utiliza para humedecer el aire.
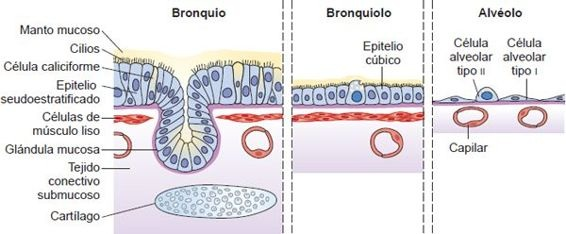
Una combinación de cartílago, fibras elásticas y de colágeno, y músculo liso da a las vías respiratorias la rigidez y flexibilidad necesarias para mantener la permeabilidad y asegurar un suministro ininterrumpido de aire. La mayor parte de las vías de conducción están revestidas con epitelio cilíndrico seudoestratificado ciliado, que contiene un mosaico de glándulas que secretan mucosidad, células ciliadas con proyecciones parecidas al cabello, y glándulas serosas que secretan un líquido acuoso que contiene enzimas bacterianas (figura 35-2). Poco a poco, la capa epitelial se vuelve más delgada cuando se mueve desde el epitelio seudoestratificado de los bronquios al epitelio cúbico de los bronquiolos y luego al epitelio escamoso de los alvéolos.
La mucosidad producida por las células epiteliales en las vías respiratorias conductoras forma una capa, llamada manto mucociliar, que protege el sistema respiratorio atrapando polvo, bacterias y otras partículas extrañas que entran a las vías respiratorias. Los cilios, que están en constante movimiento, mueven el manto mucociliar con sus partículas atrapadas como si fuese una escalera hacia la orofaringe. En este punto, el manto mucociliar se expectora o se traga. La función de los cilios en la limpieza de las vías respiratorias inferiores es óptima cuando existen niveles normales de oxígeno. La función se ve afectada cuando los niveles de oxígeno son más altos o más bajos de lo normal. Condiciones desecantes, como respirar aire caliente no humidificado durante los meses de invierno, afectan también el funcionamiento. Fumar cigarros desacelera o paraliza la motilidad de los cilios. Esta desaceleración permite que se acumulen residuos del humo del tabaco, polvo y otras partículas en los pulmones, lo que disminuye la eficiencia de este sistema de defensa pulmonar. Se cree que estos cambios contribuyen al desarrollo de bronquitis crónica y enfisema.
El agua contenida en las membranas mucosas de la vía respiratoria superior y el árbol traqueobronquial mantiene húmedas las vías respiratorias de conducción. La capacidad del aire para mantener la humedad sin condensación se incrementa cuando sube la temperatura. Así, el aire en los alvéolos, que se mantiene a la temperatura corporal, contiene por lo general una humedad mayor que el aire a temperatura atmosférica que respiramos. La diferencia entre la humedad contenida en el aire que respiramos y la que se encuentra en los alvéolos se extrae de la superficie húmeda de las membranas mucosas que revisten las vías respiratorias conductoras. Ésta es una fuente de pérdida de agua insensible. Cuando una persona tiene fiebre, el vapor de agua aumenta en los pulmones, lo que causa que se pierda más agua de la mucosa respiratoria. También, a menudo la fiebre va acompañada de un incremento en la frecuencia respiratoria, de manera que más aire que necesita hidratarse para pasar por las vías respiratorias. Como resultado, las secreciones respiratorias se engrosan, evitan el libre movimiento de los cilios y afectan la función protectora del sistema de defensa mucociliar. Esto es particularmente cierto en personas cuya ingesta de líquidos es inadecuada o en quienes experimentan deshidratación debido a otra causa patológica.
Vías respiratorias nasofaríngeas
Durante la respiración normal, la nariz es la ruta preferida para la entrada de aire hacia el tracto respiratorio. A medida que éste pasa por los pasajes nasales, se filtra, entibia y humidifica. Los pasajes lineales externos están revestidos con vellosidades gruesas, que filtran y atrapan polvo y otras partículas grandes del aire. La porción superior de la cavidad nasal está revestida con una membrana mucosa que contiene una rica red de pequeños vasos sanguíneos. Esta porción de la cavidad nasal proporciona calor y humedad al aire que respiramos.
La boca sirve como una vía respiratoria alterna cuando los pasajes nasales se taponan o cuando hay necesidad de intercambiar grandes cantidades de aire (ej. durante el ejercicio). La orofaringe se extiende posteriormente del paladar blando a la epiglotis. La orofaringe es la única abertura entre la nariz, boca y pulmones. Tanto el alimento deglutido, en su camino al esófago, como el aire, en su trayecto a la laringe, pasan por ella. La obstrucción de la orofaringe ocasiona cese inmediato de la ventilación. El control neural de la lengua y los músculos faríngeos podrían verse afectados en el coma y otros trastornos neurológicos. En estas afecciones, la lengua retrocede hacia la faringe y obstruye las vías respiratorias, en particular si la persona está recostada sobre su espalda. La hinchazón de las estructuras faríngeas causada por lesión, infección, reacción alérgica grave o la presencia de un cuerpo extraño predispone también a la obstrucción de las vías respiratorias.
Laringe
La laringe conecta la orofaringe con la tráquea. Se localiza entre la vía respiratoria superior y los pulmones. Las paredes de la laringe están soportadas por estructuras cartilaginosas firmes que evitan el colapso durante la inspiración. Las funciones de la laringe pueden dividirse en 2 categorías: las relacionadas con el habla y las que protegen a los pulmones de sustancias distintas al aire.
La cavidad de la laringe se divide en 2 pares de pliegues parecidos a estantes que se extienden del frente hacia atrás con una abertura en la línea media (figura 35-3). El par superior de pliegues, llamados pliegues vestibulares, tiene una función protectora. El par inferior de pliegues, llamados pliegues vocales, produce las vibraciones requeridas para producir sonidos vocales. Los pliegues vocales y la abertura alargada entre ellos se denominan glotis. Un conjunto complejo de músculos controla la abertura y el cierre de ésta. La epiglotis, que se localiza arriba de la laringe, es una pieza de cartílago en forma de hoja, grande, cubierta con epitelio. Cuando sólo fluye aire por la laringe, la entrada de ésta se abre y los bordes libres de la epiglotis apuntan hacia arriba. Durante la deglución, la laringe es jalada hacia arriba y los bordes libres de la epiglotis se mueven hacia abajo para cubrir la laringe, dirigiendo así los líquidos y alimentos hacia el esófago.
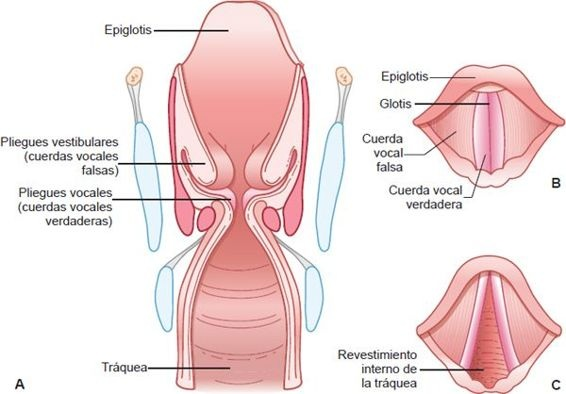
Además de la abertura y cierre de la glotis para el habla, los pliegues vocales de la laringe pueden realizar una función de esfínter, cerrando las vías respiratorias. Cuando se confrontan con sustancias distintas al aire, los músculos de la laringe se contraen y cierran la vía respiratoria. Al mismo tiempo, se inicia el reflejo tusígeno, como un medio para eliminar una sustancia extraña de la vía respiratoria. Si el mecanismo de deglución está paralizado total o parcialmente, el alimento y los líquidos pueden entrar a las vías respiratorias en lugar del esófago cuando una persona intenta deglutir. Estas sustancias no son fácilmente removidas. Cuando son jaladas hacia los pulmones, pueden causar una afección inflamatoria grave llamada neumonía por aspiración.
Árbol traqueobronquial
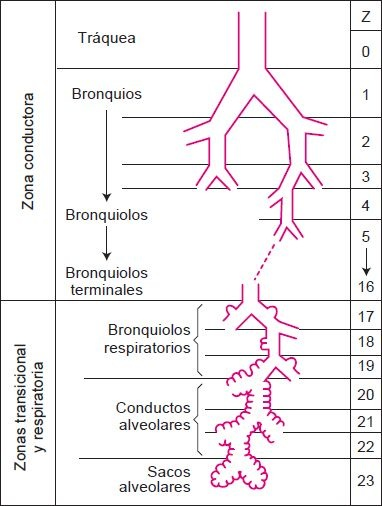
El árbol traqueobronquial, que consiste en la tráquea, bronquios y bronquiolos, puede verse como un sistema de tubos ramificados que fluyen por los lóbulos de los pulmones. Existen cerca de 23 niveles de ramificación; comienzan con las vías conductoras y terminan con las vías respiratorias, en donde tiene lugar el intercambio de gas (figura 35-4).
La tráquea es un tubo continuo que une la laringe y los bronquios mayores de los pulmones. Las paredes de la tráquea son soportadas por anillos en forma de herradura o C del cartílago hialino, que evitan que colapse cuando la presión en el tórax se vuelve negativa (figura 35-5). La parte abierta del anillo en C, que linda con el esófago, se une mediante el músculo liso. Ya que esta porción de la tráquea no es rígida, el esófago puede expandirse hacia adelante cuando el alimento deglutido pasa a través de él.
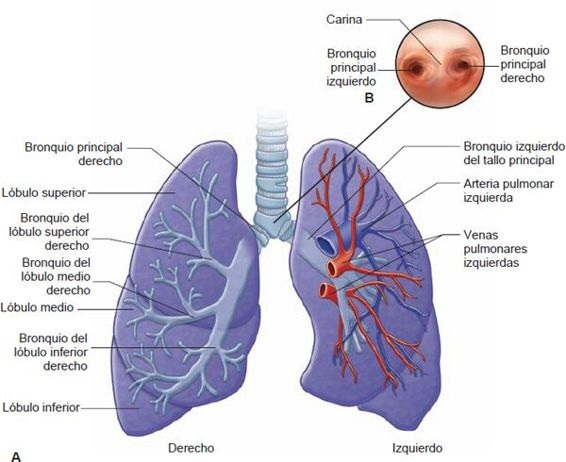
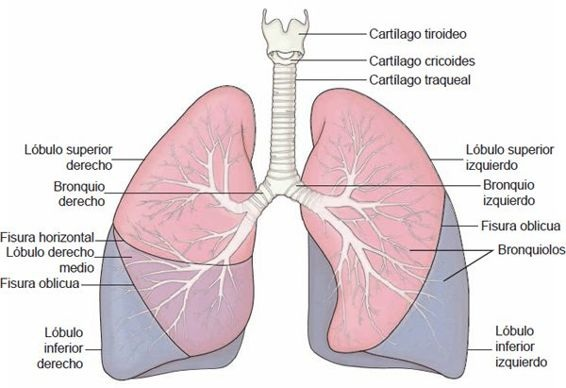
La tráquea se extiende hasta el borde superior de la quinta vértebra torácica, donde se divide para formar los bronquios principales derecho e izquierdo. Entre los bronquios principales está un borde parecido a una quilla, llamado carina (figura 35-6). La mucosa de la carina es muy sensible. Por ejemplo, la tos violenta comienza cuando un objeto extraño (ej. una uva entera, un trozo grande de hot dog o incluso la punta del catéter de succión) hace contacto con ella. La estructura de los bronquios primarios es similar a la tráquea en tanto estas vías respiratorias están revestidas con una superficie mucosa y soportadas por anillos cartilaginosos. Cada bronquio primario, acompañado de arterias pulmonares, venas y vasos linfáticos, entra al pulmón por una rendija llamada hilio.
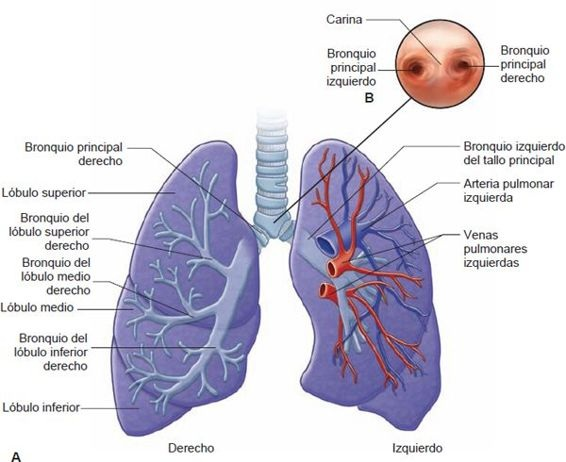
Al entrar a los pulmones, cada bronquio primario se divide en bronquios secundarios o lobulares que alimentan a cada uno de los lóbulos del pulmón, 3 en el pulmón derecho y 2 en el izquierdo (figura 35-6). El bronquio del lóbulo medio derecho es de un diámetro y longitud relativamente pequeño y, a veces, se curva de manera abrupta cerca de su bifurcación. Está rodeado por un collar de nodos linfáticos que drenan los lóbulos medio e inferior, y de manera particular está sujeto a obstrucción. A su vez, los bronquios secundarios se dividen para formar los bronquios segmentarios, que alimentan a los segmentos broncopulmonares del pulmón. La tráquea es la generación cero, los 2 bronquios del tallo principal son la primera generación y a medida que las vías respiratorias se hacen cada vez más pequeñas, se identifica su número de generación (figura 35-7).
Existen 10 segmentos en el pulmón derecho y 9 en el izquierdo; se identifican de acuerdo con su ubicación en el pulmón (ej. el segmento atípico del lóbulo superior derecho) y son las unidades más pequeñas nombradas en el pulmón. Los trastornos pulmonares como la atelectasia y la neumonía se localizan con frecuencia en un segmento broncopulmonar particular. La estructura de los bronquios secundarios y segmentarios es similar. Sin embargo, las placas irregulares de cartílago hialino que rodean por completo el lumen de los bronquios reemplazan los anillos de cartílago en forma de C. Además, hay 2 capas de músculo liso en espiral en direcciones opuestas (figura 35-8).
Los bronquios segmentarios continúan ramificándose, formando bronquios más pequeños, hasta que se convierten en bronquios terminales, la más peque ña de las vías respiratorias conductoras; a medida que se ramifican y se vuelven más pequeños, su estructura de pared cambia. El cartílago disminuye de manera gradual y hay un incremento en el músculo liso y el tejido elástico (con respecto al espesor de la pared). En el momento en que se llega a los bronquiolos no hay cartílago presente y sus paredes están compuestas sobre todo de músculo liso y fibras elásticas. El broncoespasmo, o contracción de estos músculos, causa estrechamiento de los bronquiolos e impide el flujo de aire. Las fibras elásticas, que irradian desde la capa externa de la pared bronquial y conectan con las fibras elásticas que surgen de otras partes del árbol bronquial, ejercen tensión en las paredes bronquiales. Al jalar de manera uniforme en todas direcciones, las fibras elásticas ayudan a mantener la permeabilidad de las vías respiratorias.
Pulmones y vías respiratorias
Los pulmones son las estructuras funcionales del sistema respiratorio. Además de su función de intercambio de gas, inactivan sustancias vasoactivas como bradiquinina; convierten la angiotensina I en angiotensina II y sirven como un reservorio para al almacenamiento de sangre.
Lóbulos
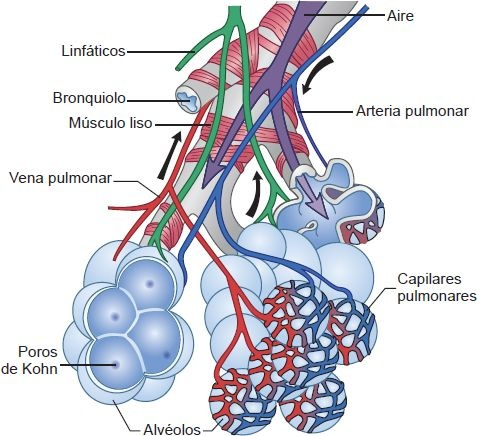
La función de intercambio de gas del pulmón tiene lugar en los lóbulos, que son las unidades funcionales más pequeñas de los pulmones. Una rama de un bronquiolo terminal, una arteriola, los capilares pulmonares y una vénula alimentan a cada lóbulo (figura 35-8).
El intercambio de gas se da en los bronquiolos respiratorios pulmonares y los ductos, y sacos alveolares. La sangre entra a los lóbulos por una arteria pulmonar y sale por una vena pulmonar. Las estructuras linfáticas rodean el lóbulo y ayudan a la remoción de proteínas plasmáticas y otras partículas de los espacios intersticiales.
A diferencia de los bronquios más grandes, los bronquiolos respiratorios están revestidos con epitelio simple en vez de epitelio seudoestratificado ciliado (figura 35-2); carecen también del soporte cartilaginoso de las vías respiratorias más grandes. En cambio, se unen al tejido elástico parecido a una esponja que contiene los espacios de aire alveolar.
Alvéolos
Los alvéolos son los espacios terminales del tracto respiratorio y los sitios reales de intercambio de gas entre el aire y la sangre. Cada alvéolo es una pequeña saculación de bronquiolos respiratorios, conductos alveolares y sacos alveolares (figura 35-8). Los sacos alveolares son estructuras de pared delgada en forma de taza que se separan entre sí por delgados tabiques alveolares. Una simple red de capilares ocupa la mayor parte de los tabiques; por tanto, la sangre está expuesta al aire en ambos lados. En el pulmón adulto existen cerca de 300 millones de alvéolos, con un área superficial total de 50 m² a 100 m² aproximadamente. A diferencia de los bronquiolos, que son tubos con sus propias paredes separadas, los alvéolos son espacios interconectados que no tienen paredes separadas. Como resultado de esta configuración, hay una mezcla continua de aire en las estructuras alveolares. Pequeños orificios en las paredes alveolares, los poros de Kohn, contribuyen también a la mezcla de aire.
El epitelio alveolar está compuesto de 2 tipos de células: células alveolares tipo I y tipo II (figura 35-9). Los alvéolos contienen células cepillo y macrófagos. Se cree que las células cepillo, que son menos en número, actúan como receptores que monitorean la calidad del aire de los pulmones, mientras que los macrófagos, presentes en el lumen alveolar y el tabique de los alvéolos, sirven para remover material problemático del pulmón.
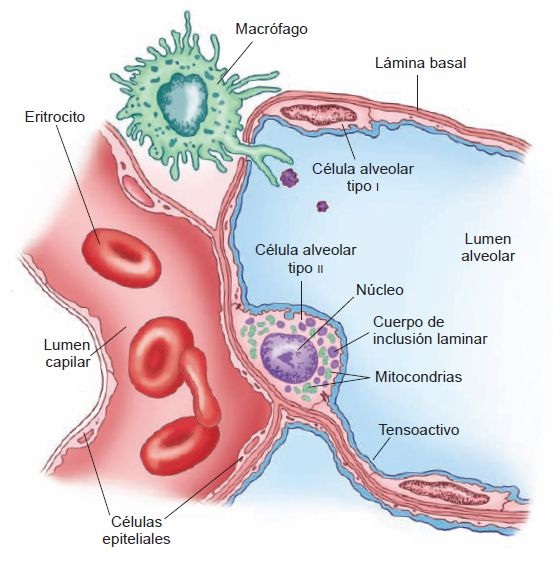
Células alveolares tipo I
Conocidas también como neumocitos tipo I, son células escamosas en extremo delgadas, con un citoplasma delgado y núcleo aplanado que ocupan cerca del 95% del área superficial de los alvéolos. Están unidas entre sí y a otras células mediante la obstrucción de uniones.
Estas uniones forman una barrera efectiva entre el aire y los componentes de la pared alveolar. Las células alveolares tipo I son incapaces de división celular.
Células alveolares tipo II
Conocidas también como neumocitos tipo II, son pequeñas células cúbicas localizadas en las esquinas de los alvéolos. Las células tipo II son tan numerosas como las células tipo I, pero, dada su forma diferente, cubren sólo cerca del l5% del área superficial alveolar. Las células tipo II sintetizan tensoactivo pulmonar, una sustancia que disminuye la tensión superficial en los alvéolos y propicia una mayor facilidad de inflación pulmonar. Son también células progenitoras para las células tipo I. Después de la lesión pulmonar, proliferan y restauran las células alveolares tipo I y tipo II.
El tensoactivo pulmonar es una mezcla compleja de fosfolípidos, lípidos neutros y proteínas que se sintetizan en las células alveolares tipo II. Estas últimas son ricas en mitocondrias y, además, metabólicamente activas. Su citoplasma atípico contiene pilas de láminas de membrana paralelas o laminillas, llamadas cuerpos laminares. La secreción de tensoactivo tiene lugar por exocitosis.
Dentro del pulmón, la principal ruta de eliminación de tensoactivo es a través de la recaptación mediante las células tipo II. Después de la recaptación, los fosfolípidos son reciclados o degradados y reutilizados en la síntesis de nuevos fosfolípidos.
Las moléculas de tensoactivo producidas por las células alveolares tipo II reducen la tensión superficial en la interfase aire-epitelio y modulan las funciones inmunes del pulmón. Existen 4 tipos de tensoactivo, cada uno con una estructura molecular diferente: apoproteínas tensoactivas A (SP-A), B (SP-B), C (SPC) y D (SP-D).
SP-B y SP-C reducen la tensión superficial en la interfase aire-epitelio e incrementan la distensibilidad pulmonar y facilidad de inflación del pulmón. SP-B es en especial importante para la generación de la película reductora de superficie que hace posible la expansión del pulmón. Por ejemplo, los trastornos por insuficiencia de SP-B son con frecuencia resultado de trastornos por un solo gen que causan disfunción respiratoria aguda o crónica 2. SP-C es necesaria para disminuir la tensión superficial y detener el colapso alveolar expiratorio terminal 3.
SP-A y SP-D no reducen la tensión superficial, pero contribuyen con las defensas huésped que protegen contra patógenos que han entrado al pulmón. Son los miembros de la familia de la proteína colectina, que funciona como una parte del sistema inmune congénito. En conjunto opsonizan patógenos, incluso bacterias y virus, para facilitar la fagocitosis mediante macrófagos. Regulan también la producción de mediadores inflamatorios. SP-A y SP-D son directamente bactericidas, lo que significa que pueden matar bacterias en ausencia de células efectoras del sistema inmune. SP-D pueden utilizarse para predecir problemas con la lesión pulmonar aguda (LPA) y el síndrome de dificultad respiratoria aguda (SDRA). Ambas afecciones pueden predecirse mejor mediante una combinación de biomarcadores (SP-D, factor quimiotáctico de neutrófilos e interleucina-8) y predictores clínicos (Acute Physiology and Cronic Health Evaluation III -APACHE III-). Por consiguiente, resulta más significativo medir SP-D en personas que experimentan traumatismo torácico.
Macrófagos alveolares
Los macrófagos están presentes en el tejido conectivo del tabique y en los espacios de aire de los alvéolos. Son responsables de la remoción de sustancias problemáticas de los alvéolos. En los espacios de aire barren la superficie para remover partículas inhaladas, como polvo y polen. Algunos macrófagos pasan al árbol bronquial en la mucosa y se eliminan mediante deglución o tosiendo cuando llegan a la faringe. Otros entran al tejido conectivo septal, donde, llenos de material fagocitado, permanecen de por vida. Así, en la autopsia, los residentes urbanos, así como los fumadores, muestran a menudo muchos macrófagos alveolares llenos de carbono y otras partículas contaminantes del ambiente.
Los macrófagos alveolares fagocitan también agentes infecciosos insolubles, como Mycobacterium tuberculosis. Los macrófagos activados se agregan entonces para formar un granuloma encapsulado en fibrina, llamado tubérculo, para contener la infección. El bacilo tuberculoso puede permanecer inactivo en esta etapa o ser reactivado años después, cuando la tolerancia inmunitaria de la persona disminuye como resultado de la edad avanzada, enfermedad o tratamiento inmunosupresor.
Vasculatura pulmonar y suministro linfático
Circulación pulmonar y bronquial
Los pulmones se abastecen con una alimentación sanguínea dual, las circulaciones pulmonares y bronquial. La circulación pulmonar surge de la arteria pulmonar y proporciona la función de intercambio de gas de los pulmones (figura 35-8). La sangre desoxigenada sale del corazón derecho por la arteria pulmonar. La arteria pulmonar se divide en una arteria pulmonar izquierda que entra al pulmón izquierdo, y una arteria pulmonar derecha que entra al pulmón derecho. El regreso de la sangre oxigenada al corazón se da mediante las venas pulmonares, que desembocan en la aurícula izquierda. Ésta es la única parte de la circulación en donde las arterias llevan sangre desoxigenada y las venas llevan sangre oxigenada.
La circulación bronquial distribuye sangre a las vías respiratorias conductoras y las estructuras de soporte del pulmón; tiene también una función secundaria: calentar y humidificar el aire entrante a medida que se mueve por las vías respiratorias conductoras. Las arterias bronquiales salen de la aorta torácica y entran a los pulmones con los bronquios mayores. Se dividen y subdividen junto con los bronquios a medida que se salen hacia los pulmones, proporcionando oxígeno a éstos y otras estructuras pulmonares. La sangre de los capilares en la circulación bronquial drena hacia las venas bronquiales, donde la sangre de las venas bronquiales más grandes desembocan en la vena cava. La sangre de las venas bronquiales más pequeñas se vacía en las venas pulmonares. Dado que la circulación bronquial no participa en el intercambio de gas, esta sangre se desoxigena. Como resultado, diluye la sangre oxigenada y vuelve al lado izquierdo del corazón por las venas pulmonares.
Los vasos sanguíneos bronquiales son los únicos que pueden experimentar angiogénesis (formación de nuevos vasos) y desarrollar circulación colateral cuando se obstruyen los vasos en la circulación pulmonar, como en el embolismo pulmonar. El desarrollo de nuevos vasos sanguíneos ayuda a mantener vivo el tejido pulmonar hasta que se restablece la circulación pulmonar.
Circulación linfática
Los pulmones son abastecidos también con drenaje linfático, que es análogo a su suministro sanguíneo dual. Un conjunto de vasos linfáticos, los vasos superficiales, drena la superficie del pulmón y viaja en el tejido conectivo de la pleura visceral. Un segundo conjunto de vasos, los vasos linfáticos profundos, sigue las arterias pulmonares, venas pulmonares y árbol bronquial hacia abajo hasta el nivel de los bronquiolos respiratorios (figura 35-8). Ambos sistemas tienen numerosas interconexiones y forman redes que drenan hacia los nodos linfáticos hiliares en la base de cada pulmón. Las partículas que entran al pulmón se eliminan en parte en estos canales, como las proteínas plasmáticas que han escapado de los capilares pulmonares. Esta última función es en especial importante para mantener secos los pulmones y evitar la acumulación de líquido en la cavidad pleural.
Inervación
Las divisiones simpáticas y parasimpáticas del sistema nervioso autónomo inervan el pulmón. Su estimulación parasimpática, a través del nervio vago, se encarga del tono del músculo liso ligeramente restringido en el pulmón normal en reposo. No hay inervación motora voluntaria del pulmón ni fibras de dolor. Las fibras de dolor se encuentran sólo en la pleura.
La estimulación del sistema nervioso parasimpático origina la constricción de las vías respiratorias y el incremento de la secreción glandular. La inervación parasimpática del pulmón surge de los núcleos vagales en la médula. Las fibras preganglionares de los núcleos vagales descienden en el nervio vago hacia ganglios adyacentes a las vías respiratorias y vasos sanguíneos del pulmón. Las fibras posganglionares de los ganglios completan la red que inerva el músculo liso, vasos sanguíneos y células epiteliales, incluso las células caliciformes y glándulas submucosas. Tanto las fibras preganglionares como las posganglionares contienen neuronas motoras excitatorias (colinérgicas) que responden a acetilcolina. La inervación parasimpática es mayor en las vías respiratorias grandes y disminuye hacia las vías respiratorias más pequeñas.
La estimulación del sistema nervioso simpático causa relajación de las vías respiratorias, constricción de vasos sanguíneos e inhibición de secreción glandular. La inervación simpática surge de los cuerpos celulares en los ganglios simpáticos paraverterbrales. Los neurotransmisores del sistema nervioso simpático incluyen las catecolaminas noradrenalina y adrenalina.
Pleura
Una membrana serosa, delgada, transparente, de doble capa, llamada pleura, reviste la cavidad torácica y encierra a los pulmones (figura 35-10). La capa parietal externa cubre las cavidades pulmonares y se adhiere a la pared torácica, el mediastino y el diafragma. La pleura visceral interna cubre estrechamente el pulmón y se adhiere a toda su superficie. Continúa en la pleura parietal y el hilio del pulmón, en donde el bronquio mayor y los vasos pulmonares entran y salen del pulmón.
Una delgada película de líquido seroso separa las 2 capas pleurales, lo que permite que ambas capas se deslicen entre sí pero que se mantengan juntas. De este modo, no hay separación entre los pulmones y la pared torácica, que potencialmente permitiría que se acumulara el proceso infeccioso y afectara de modo negativo la expansión de la pared torácica. La cavidad pleural es un espacio potencial donde se puede acumular el líquido seroso o exudado inflamatorio. El término efusión pleural se emplea para describir una acumulación anómala de líquido o exudado en la cavidad pleural.