01. Sistema de conducción cardíaco
En ciertas áreas del corazón, las células miocárdicas se modifican para convertirse en células especializadas del sistema de conducción. Estas células especializadas tienen la capacidad de iniciar y conducir impulsos. El sistema de conducción es el que mantiene la eficiencia de bombeo del corazón. Las células marcapasos especializadas generan impulsos a una frecuencia más rápido que las células de otros tipos de tejido cardíaco y el tejido de conducción transmite estos impulsos a mayor velocidad que otros tipos de células cardíacas. Gracias a estas propiedades, un sistema de conducción normal controla la frecuencia del corazón.
El sistema especializado estimulante y de conducción del corazón consiste en el nodo senoauricular (SA), en el que se genera un impulso rítmico; las vías internodales entre las aurículas y los ventrículos; el nodo auriculoventricular (AV) y el haz de His, que conduce el impulso de las aurículas a los ventrículos; y las fibras de Purkinje, que conducen los impulsos por todo el tejido de los ventrículos derecho e izquierdo (figura 33-1).
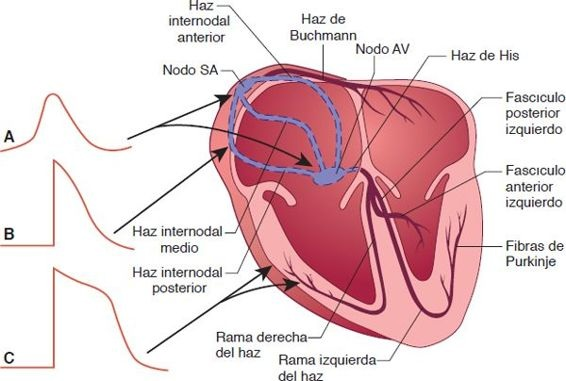
El corazón tiene 2 sistemas de conducción: uno que controla la actividad auricular y uno que controla la actividad ventricular. La conducción auricular comienza en el nodo SA, que tiene la frecuencia intrínseca de disparo más rápido (60 lpm a 100 lpm) y actúa como el marcapasos normal del corazón. Es una franja fusiforme de tejido muscular especializado de 10 mm a 20 mm de largo y de 2 mm a 3 mm de ancho, situada en la pared posterior de la aurícula derecha, justo debajo de la abertura de la vena cava superior y a menos de 1 mm de la superficie epicárdica. Se ha sugerido que no hay una célula individual en el nodo SA que funcione como marcapasos, sino que las células nodales sinusales emiten descargas sincrónicas por la estimulación mutua. Como resultado, el disparo de las células de emisiones más rápidas se enlentece por las de descarga más lenta y la frecuencia de disparo de las células lentas se acelera por influencia de las células rápidas, lo que produce la sincronización de sus frecuencias de disparo y esto no permite que la conducción de las aurículas a los ventrículos sea demasiado rápida.
Los impulsos que se originan en el nodo SA se transmiten por las aurículas al nodo AV. Debido a la localización anatómica del nodo SA, la progresión de la despolarización auricular sigue una dirección inferior, izquierda y un poco posterior, y la aurícula derecha se despolariza un poco antes que la izquierda. Existen 3 vías internodales entre el nodo SA y el nodo AV: los haces anterior (de Bachmann), medio (de Wenckebach) y posterior (de Thorel). Estos 3 haces se unen antes de llegar al nodo AV. Este haz muscular grande se origina a lo largo del borde anterior del nodo SA y transcurre hacia atrás, alrededor de la aorta hacia la aurícula izquierda.
La unión AV conecta los 2 sistemas de conducción y establece una conducción unidireccional entre las aurículas y los ventrículos.
El nodo AV es una estructura ovoide compacta que mide alrededor de 1 mm × 3 mm × 5 mm y se localiza en la pared posterior, un poco debajo del endocardio auricular derecho, anterior a la abertura del seno coronario y justo por arriba de la inserción de la valva septal de la válvula tricúspide. Es importante señalar que en todo el corazón sano, excepto el nodo AV, el músculo auricular está separado del músculo ventricular para impedir el disparo de impulsos cardíacos inadecuados.
El nodo AV se divide en 3 regiones funcionales:
- La región AN o de transición, situada entre las aurículas y el resto del nodo.
- La región N o medial (o sea, el propio nodo).
- La región NH, en la que las fibras nodales se fusionan con el haz de His, que es la porción superior del sistema especializado de conducción.
En la porción AN del nodo, las fibras auriculares se conectan con fibras de la unión muy pequeñas del nodo mismo. La velocidad de conducción por las fibras AN y N es muy baja (casi la mitad de la del músculo cardíaco normal), lo que retrasa mucho la transmisión del impulso. Existe un retraso adicional cuando el impulso se transmite por la región N hacia la región NH, que se conecta con el haz de His (también llamado haz AV). Este retraso proporciona una ventaja mecánica, ya que las aurículas completan la expulsión de sangre antes que comience la contracción ventricular. En condiciones normales, el nodo AV es la única conexión entre los sistemas de conducción auricular y ventricular. Su principal trabajo es coordinar la conducción auricular y ventricular. Si se bloqueara la transmisión de los impulsos por el nodo AV, las aurículas y los ventrículos se contraerían de manera independiente.
El sistema de Purkinje, que inicia la conducción ventricular, tiene fibras grandes que permiten laconducción rápida. Una vez que el impulso entra al sistema de Purkinje, se extiende casi de inmediato al ventrículo completo (0,03 s). Esta rapidez de conducción por todo el sistema de Purkinje es necesaria para la expulsión rápida y eficiente de sangre del corazón. Las fibras del sistema de Purkinje se originan en el nodo AV y luego forman el haz de His, que se extiende por el tejido fibroso entre las válvulas cardíacas y en el sistema ventricular. Debido a su proximidad con la válvula aórtica y el anillo valvular mitral, el haz de His está predispuesto a la inflamación y depósito de detritos calcificados que pueden interferir con la conducción del impulso. El haz de His penetra en las ramas derecha e izquierda que se extienden a ambos lados del tabique interventricular. Las ramas de las arterias coronarias descendentes anterior y posterior irrigan el haz de His, lo que vuelve su conducción menos susceptible al daño isquémico, a menos que el daño sea excesivo. Las ramas del haz se distribuyen por los tejidos subendocárdicos hacia los músculos papilares y luego se subdividen en fibras de Purkinje, que se ramifican y se distribuyen por las paredes externas de los ventrículos. El tronco principal de la rama izquierda se extiende 1 cm o 2 cm antes de dividirse en múltiples ramas a su entrada al área septal para separarse además en 2 segmentos: los fascículos izquierdos posterior y anterior. En el sistema de conducción sano, estas fibras de Purkinje transmiten el impulso casi al mismo tiempo al endocardio ventricular derecho e izquierdo.
Cuando las fibras nodales AV no se estimulan, emiten descargas una frecuencia intrínseca de 40 lpm a 60 lpm, las fibras de Purkinje emiten de 15 a 40 descargas por min. Aunque el nodo AV y el sistema de Purkinje tienen la capacidad de controlar la frecuencia del corazón, en condiciones normales no lo hacen porque la frecuencia de descarga del nodo SA es mucho más rápida. Cada vez que el nodo SA emite una descarga, el impulso se conduce al nodo AV y las fibras de Purkinje, y hace que éstos emitan una descarga. El nodo AV puede asumir la función de marcapasos del corazón en caso que el nodo SA no emita descargas y el sistema de Purkinje puede asumir la función de marcapasos ventricular si el nodo AV no conduce impulsos de las aurículas a los ventrículos. En estas circunstancias, la frecuencia cardíaca también refleja la frecuencia intrínseca de descarga de las estructuras prevalecientes.
Potenciales de acción
Un potencial de acción es un cambio secuencial en el potencial eléctrico que ocurre a través de una membrana celular cuando se produce la excitación que se conduce por las aurículas y los ventrículos.
Estos potenciales o diferencias de voltaje, a menudo llamados potenciales de membrana, representan el flujo de corriente producido por el paso de iones a través de conductos iónicos en la membrana celular. Los iones sodio (Na+), potasio (K+) y calcio (Ca2+) son los principales portadores de carga en las células miocárdicas. Cada vez se considera más probable que los trastornos en los conductos iónicos y la interrupción del flujo de estos iones portadores de corriente estén vinculados con el origen de las arritmias cardíacas y los trastornos de la conducción.
Los potenciales de acción pueden dividirse en 3 fases:
- Estado en reposo o no excitado.
- Despolarización.
- Repolarización.
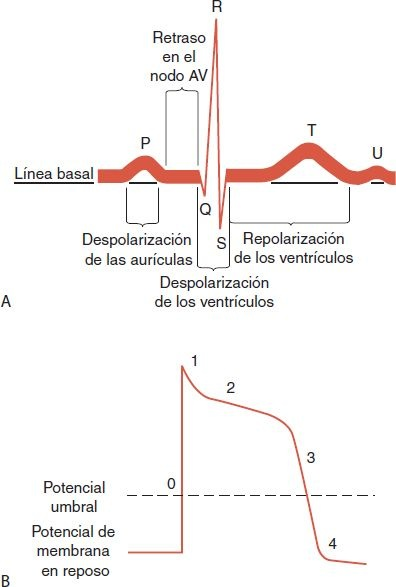
Durante la fase de reposo, las células cardíacas mantienen un potencial de membrana en reposo que varía entre −60 mV y −90 mV. El signo negativo previo al voltaje indica que el interior de la membrana tiene carga negativa en relación con el exterior (figura 33-2A). Aunque existen distintos tipos de iones en el interior y el exterior de la membrana, el potencial de membrana depende sobre todo del Na+ y el K+, así como de la permeabilidad de la membrana a éstos iones. Durante la fase de reposo del potencial de membrana, ésta mantiene permeabilidad selectiva al K+ y es casi impermeable al Na+. Como resultado, el K+ difunde fuera de la célula en favor de su gradiente de concentración, lo que causa una pérdida relativa de iones positivos del interior de la membrana. El resultado es una distribución desigual de la carga, con la negatividad en el interior y la positividad en el exterior.

La despolarización es el período (medido en milisegundos [ms]) durante el cual se revierte la polaridad del potencial de membrana. Ocurre cuando la membrana celular se vuelve selectivamente permeable de manera súbita a un ion portador de corriente, como el Na+, lo que permite su entrada a la célula y cambia el potencial de membrana, que se vuelve positivo en el interior y negativo en el exterior (figura 33-2B).
La repolarización es el restablecimiento del potencial de membrana en reposo. Es un proceso complejo y algo lento, incluye el flujo de salida de cargas eléctricas de la célula y el regreso del potencial de membrana a su estado de reposo. Durante la repolarización, la permeabilidad de la membrana para K+ aumenta de nuevo, lo que permite que el K+ con carga positiva salga a través de la membrana. Este desplazamiento hacia el exterior elimina cargas positivas del interior de la célula; por tanto, el voltaje a través de la membrana se vuelve negativo de nuevo en el interior y positivo en el exterior (figura 33-2C). La bomba de sodio-potasio dependiente de trifosfatasa de adenosina (ATPasa) ayuda a la repolarización mediante el bombeo de las cargas positivas hacia fuera a través de la membrana celular y el regreso de K+ hacia el interior de la membrana.
Fases del potencial de acción
Los potenciales de acción en el músculo cardíaco casi siempre se dividen en 5 fases:
- Fase 0: ascenso o despolarización rápida.
- Fase 1: período de repolarización rápida.
- Fase 2: meseta
- Fase 3: período de repolarización final rápida.
- Fase 4: despolarización diastólica (figura 33-3B).
El músculo cardíaco tiene 3 tipos de conductos iónicos de membrana que contribuyen a los cambios en el voltaje que ocurren durante las distintas fases del potencial de acción cardíaco. Son los conductos rápidos de Na+, conductos lentos de calcio (Na2+) y conductos de K+.
Durante la fase 0, se estimula a los conductos de Na+ de la membrana celular para abrirse, lo que permite la entrada rápida de Na+ ; esto ocurre en el músculo auricular y ventricular, y en las fibras de Purkinje. El punto en que los conductos de Na+ se abren se llama umbral de despolarización. Cuando la célula llega a este umbral, se produce la entrada rápida de Na+. El exterior de la célula queda con carga negativa en relación con el interior celular muy positivo. Esta entrada de Na+ produce un cambio rápido en el potencial de membrana dirigido a la polaridad positiva, lo que genera la espiga eléctrica y ascenso excesivo durante la fase 0 del potencial de acción. El potencial de membrana cambia desde un valor cercano a 90 mV hasta +20 mV. La despolarización rápida que comprende la fase 0 es la causa del complejo QRS en la electrocardiografía (ECG) (figura 33-3A).
La despolarización de una célula cardíaca tiende a producir despolarización de las células adyacentes porque la espiga de voltaje de la célula que se despolariza abre los conductos de Na+ en las células cercanas. Por lo tanto, cuando una célula cardíaca se estimula y se despolariza, se propaga una onda de despolarización por el corazón, célula por célula.
La fase 1 ocurre en el pico del potencial de acción y señala la desactivación de los conductos rápidos de Na+ con un descenso súbito en la permeabilidad al sodio. Se cree que la ligera pendiente descendente se debe a la entrada de una pequeña cantidad de iones cloruro con carga negativa y salida de potasio. La disminución de la positividad intracelular reduce el potencial de membrana a un nivel cercano a 0 mV, a partir del cual surge la meseta, o fase 2.
La fase 2 representa la meseta del potencial de acción. Si la permeabilidad al K+ aumentara hasta su nivel en reposo para este momento, como sucede en las fibras nerviosas o el músculo esquelético, la célula se repolarizaría con rapidez. En lugar de eso, la permeabilidad al K+ es baja, lo que permite que la membrana se mantenga despolarizada durante toda la meseta de la fase 2. La entrada concurrente de Ca2+ a la célula por los conductos lentos de Ca2+ contribuye a la meseta de la fase 2. Los iones calcio que entran al músculo durante esta fase también tienen un papel clave en el proceso contráctil. Estas características únicas de la meseta fase 2 generan el potencial de acción del músculo cardíaco (varios cientos de milisegundos), que dura 3 a 15 veces más que el del músculo esquelético y produce una contracción más prolongada, en consecuencia. La meseta de la fase 2 coincide con el segmento ST de la ECG.
La fase 3 refleja la repolarización rápida y comienza con el descenso del potencial de acción. Durante el período de repolarización de la fase 3, los conductos lentos de Ca2+ se cierran y la entrada de Ca2+ y Na+ cesa. Existe un aumento agudo en la permeabilidad al K+, lo que contribuye al desplazamiento rápido de K+ al exterior y restablecimiento del potencial de membrana en reposo (-90 mV). Al final de la fase 3, la distribución de K+ y Na+ normaliza el estado de reposo de la membrana. La onda T de la ECG corresponde a la fase 3 del potencial de acción.
La fase 4 representa el potencial de membrana en reposo. Durante esta fase, la actividad de la bomba de Na+/K+-ATPasa contribuye al mantenimiento del potencial de membrana en reposo mediante el transporte de Na+ fuera de la célula y el regreso del K+ hacia el interior. La fase 4 corresponde a la diástole.
Respuestas rápida y lenta
Existen 2 tipos principales de potenciales de acción en el corazón: la respuesta rápida y la respuesta lenta.
La respuesta rápida ocurre en las células miocárdicas normales de las aurículas, los ventrículos y las fibras de Purkinje (figura 33-4A). Se caracteriza por la abertura de conductos de Na+ dependiente de voltaje llamados conductos rápidos de sodio. Las células cardíacas de respuesta rápida no inician potenciales de acción en condiciones normales.
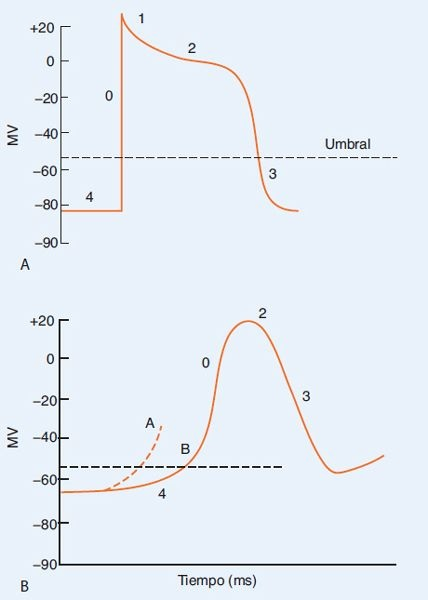
Las células de respuesta rápida tienen un potencial de reposo constante, despolarización rápida y luego un período más largo de despolarización sostenida antes de la repolarización. Esto permite la conducción rápida del impulso a las células adyacentes. Las fibras miocárdicas con respuesta rápida son capaces de conducir actividad eléctrica a frecuncias relativamente rápidas (0,5 m/s a 5,0 m/s), lo que representa un factor de alta seguridad para la conducción.
La respuesta lenta ocurre en el nodo SA, que es el marcapasos natural del corazón, y las fibras de conducción del nodo AV (figura 33-4B). La característica distintiva de estas células marcapasos es la despolarización espontánea en la fase 4. La permeabilidad de la membrana de estas células permite el escape lento al interior de la corriente a través de los conductos lentos durante la fase 4. Esta fuga continúa hasta que se alcanza el umbral para la descarga, momento en el cual la célula se despolariza de manera espontánea. En condiciones normales, la respuesta lenta, a veces llamada corriente de calcio, no contribuye mucho a la despolarización de las aurículas y los ventrículos. Su papel principal en las células auriculares y ventriculares normales es permitir la entrada de calcio para el mecanismo de excitacióncontracción que acopla la actividad eléctrica con la contracción muscular.
La velocidad con la que las células marcapasos emiten descargas varía con el potencial de membrana en reposo y la pendiente de la despolarización de la fase 4 (figura 33-3). Las catecolaminas (adrenalina y noradrenalina) elevan la frecuencia cardíaca porque aumentan la pendiente o la frecuencia de despolarización en la fase 4. La acetilcolina, un mediador parasimpático, reduce la frecuencia cardíaca porque disminuye la pendiente de la fase 4.
La respuesta rápida del músculo auricular y ventricular puede convertirse en una respuesta de marcapasos lenta en ciertas condiciones. Por ejemplo, esta conversión puede ocurrir de manera espontánea en sujetos con enfermedad coronaria grave, en áreas del corazón en las que el suministro sanguíneo está muy reducido. Los impulsos generados por estas células pueden generar latidos ectópicos y arritmias graves.
Períodos refractarios absolutos y relativos
La acción de bombeo del corazón requiere la contracción y relajación alternadas. Existe un período en el potencial de acción durante el cual la membrana no puede estimularse para generar otro potencial de acción (figura 33-5). Este período, llamado período refractario absoluto o efectivo, incluye las fases 0, 1 y 2, y parte de la fase 3. Durante este intervalo, la célula no puede despolarizarse de nuevo en ninguna circunstancia. Esto actúa como margen de seguridad cardíaca a fin de prevenir cualquier estímulo o la generación de latidos adicionales. Cuando la repolarización regresó el potencial de membrana por debajo del umbral, aunque aún no sea el potencial de membrana en reposo (−90 mV), la célula es capaz de responder a un estímulo mayor de lo normal. Esta condición se conoce como período refractario relativo. El período refractario relativo comienza cuando el potencial de membrana en la fase 3 alcanza el umbral y termina justo antes del final de la fase 3.
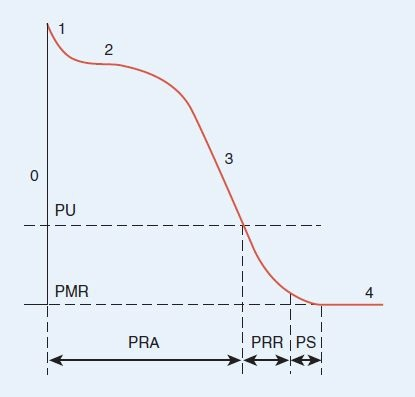
Después del período refractario relativo existe un intervalo corto llamado período excitatorio supranormal, durante el cual un estímulo débil puede inducir una respuesta. Este período va desde la parte final de la fase 3 hasta el inicio de la fase 4. Durante ese período es que se desarrollan las arritmias cardíacas.
En el músculo esquelético, el período refractario de es muy corto con respecto a la contracción, por lo que puede iniciarse una contracción antes que termine la primera y esto produce una contracción tetánica sumada. En el músculo cardíaco, el período refractario absoluto es casi tan largo como la contracción misma, lo que asegura que no pueda iniciarse una segunda contracción hasta que la primera se complete. La mayor duración del período refractario absoluto es importante paramantener la contracción alternada con la relajación, esencial para la acción de bombeo del corazón y para prevenir las arritmias mortales.
Electrocardiografía
La ECG es un registro gráfico de la actividad eléctrica del corazón o una imagen del corazón mientras se contrae. La corriente eléctrica generada por el corazón se extiende por el cuerpo hasta la piel, donde puede percibirse con electrodos en sitios apropiados, amplificarse y presentarse en un osciloscopio o registro gráfico.
Los puntos de desviación en una ECG se designan con las letras P, Q, R, S, y T. La onda P representa la despolarización del nodo SA y las aurículas; el complejo QRS (o sea, del inicio de la onda Q al final de la onda S) muestra la despolarización ventricular; y la onda T representa la repolarización ventricular. La línea isoeléctrica entre la onda P y la onda Q representa la despolarización del nodo AV, las ramas y el sistema de Purkinje. La repolarización auricular ocurre durante la despolarización ventricular y queda oculta en el complejo QRS. La figura 33-6 muestra la actividad eléctrica del sistema de conducción en un trazo ECG.
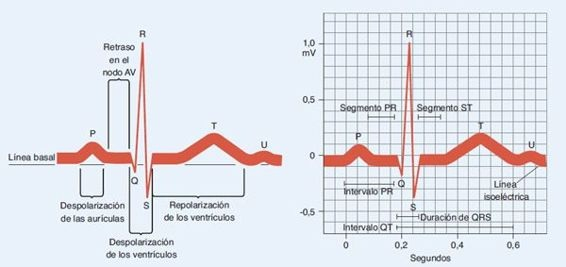
La ECG registra la diferencia potencial en la carga entre 2 electrodos conforme las ondas de despolarización y repolarización avanzan por el corazón y se conducen a la superficie de la piel. La forma del trazo en el registro depende de la dirección en la que el impulso se propaga por el músculo cardíaco en relación con el sitio del electrodo. Una onda de despolarización que se mueve hacia el electrodo de registro se marca como una desviación positiva, ascendente. Por el contrario, si el impulso se propaga en sentido contrario al electrodo de registro, la desviación es hacia abajo o negativa. Cuando no hay flujo o cambio entre los electrodos, el potencial es cero y se registra una línea recta en la línea basal de la gráfica.
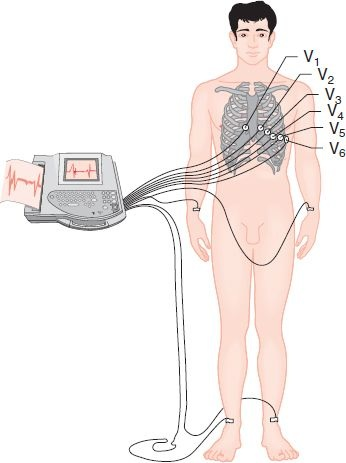
Por convención se obtiene el registro en 12 derivaciones (6 derivaciones en extremidades y 6 derivaciones torácicas) para la ECG diagnóstica; cada una proporciona una vista única de las fuerzas eléctricas del corazón desde una posición distinta en la superficie del cuerpo. Las 6 derivaciones de las extremidades perciben las corrientes eléctricas a su paso por el corazón en el plano frontal o vertical. Los electrodos se conectan con las 4 extremidades o zonas representativas en el cuerpo cerca de los hombros y la parte inferior torácica o abdominal. El potencial eléctrico registrado en cualquier extremidad debe ser el mismo, sin importar el punto en la extremidad donde se coloque el electrodo. Las 6 derivaciones torácicas proporcionan una vista de las corrientes eléctricas a su paso por el corazón en el plano horizontal. Se mueven a distintas posiciones en el pecho, incluidos los bordes esternales derecho e izquierdo y la superficie anterior izquierda (figura 33-7). La derivación de la extremidad inferior derecha se usa como electrodo de tierra. Cuando está indicado, pueden aplicarse más electrodos en otras áreas del cuerpo, como la espalda o la región anterior derecha del tórax.
Aunque la colocación exacta y la selección de las derivaciones son aspectos importantes en la vigilancia ECG, hay muchos estudios que identificaron 2 errores frecuentes: colocación inexacta de los electrodos y selección incorrecta de las derivaciones para situaciones clínicas específicas. La colocación incorrecta de los electrodos puede cambiar mucho la morfología del complejo QRS, lo que conduciría a un diagnóstico incorrecto de arritmias cardíacas o falta de detección de defectos en la conducción existentes. También puede hacer que las personas reciban tratamientos inadecuados que pueden ser dañinos.
En pacientes con síndrome coronario agudo (SCA), que incluye angina inestable e infarto de miocardio (IM) con o sin elevación del segmento ST, la vigilancia ECG cuidadosa es obligada. Las personas con SCA tienen riesgo de que se extienda el área infartada, isquemia de miocardio constante y arritmias que ponen en peligro su vida. La investigación mostró que la vigilancia ECG es más sensible que el informe de síntomas del sujeto para identificar la isquemia de miocardio transitoria.
La vigilancia ECG también permite la detección más exacta y oportuna de fenómenos isquémicos que predicen complicaciones tempranas. Asimismo, la vigilancia ECG es esencial para decidir las opciones terapéuticas, como la reperfusión. Se recomienda usar las 12 derivaciones ECG para vigilar a los pacientes con SCA porque los cambios isquémicos pueden ser evidentes en distintas derivaciones en diferentes momentos.
En fecha reciente, la American Heart Association publicó los estándares prácticos para la vigilancia ECG en el hospital. Este sistema de calificación incluye 3 categorías:
- Clase I : la vigilancia cardíaca es necesaria en la mayoría, si no en todas las personas de este grupo.
- Clase II : la vigilancia cardíaca puede ser provechosa en algunas personas, pero no es un elemento esencial de la atención de estos pacientes.
- Clase III : la vigilancia cardíaca no está indicada porque el riesgo de un incidente adverso en estas personas es tan bajo que la vigilancia no se considera terapéutica.
Los ejemplos de pacientes incluidos en la clase I son los que se reanimaron de un paro cardíaco; que están en la fase temprana de un SCA; tienen un síndrome coronario inestable o lesiones coronarias de alto riesgo recién diagnosticadas; o que se sometieron a cirugía cardíaca en las 48 h a 72 h previas.
Además, se incluyen recomendaciones para asignación de personal, entrenamiento, documentación y estrategias para mejorar la calidad de la vigilancia ECG. Se recomienda seguir los estándares para la práctica cuando se toman decisiones sobre la vigilancia ECG.