Reproducción Humana
Anatomía de los órganos reproductores femeninos.
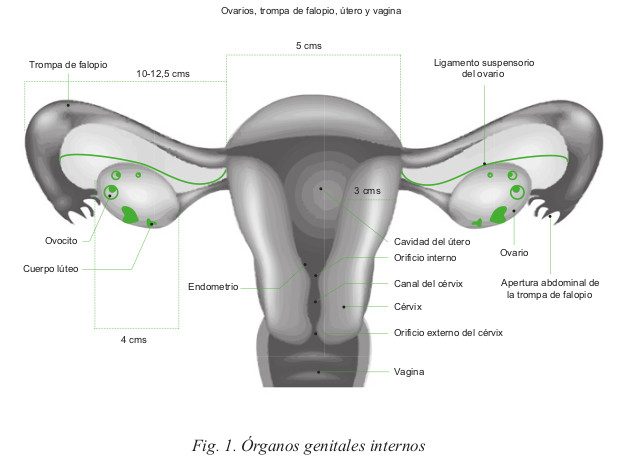
Órganos genitales internos
En donde distinguimos, entre otras, las siguientes estructuras:
A. Ovarios
El ovario es un órgano intrapélvico que desempeña dos funciones primordiales: producción del gameto femenino y secreción de hormonas esteroideas.
Es un órgano par, tienen una forma ovoidea que algunos autores comparan con la forma de una almendra. Sus dimensiones cambian a lo largo de la vida de la mujer entre 30-40 mm de longitud, 20 mm de anchura y 10-15 mm de espesor.
Su coloración es blanca rosada en el sujeto vivo y gris blanquecina en el sujeto cadáver. Su superficie es lisa antes de iniciarse la pubertad, a partir de este momento presentan una serie de eminencias producidas por las formaciones foliculares.
Cada ovario está unido a la superficie posterior del ligamento ancho del útero, a través de un pliegue de peritoneo denominado mesoovario.
La parte superior del ovario está fijada mediante el ligamento lumboovárico a la pared de la pelvis. Por este ligamento también viajan los nervios y vasos sanguíneos que inervan e irrigan el ovario. La parte inferior está unida al útero mediante el ligamento ovárico.
a. Constitución del ovario
En un corte transversal distinguimos, desde exterior a interior, varias zonas en el ovario:
- Epitelio ovárico: es una capa de células cuboidales que recubre el ovario.
- Túnica albugínea: es una capa de tejido conjuntivo denso fibroso con células fusiformes. Es una capa más densa que la que recubre los testículos y su función es eminentemente protectora.
- La corteza: está formada por el estroma, constituida por tejido similar al conectivo pero con una gran cantidad de células, muy próximas unas a otras, respecto a la matriz extracelular, que posee pocas fibras reticulares y de colágeno. Estas células tienen como misión principal producir hormonas esteroideas. El estroma está altamente vascularizado. Entre las células del estroma se encuentran los folículos en diferentes estados de maduración: folículos primordiales, folículos en maduración, folículos de Graaf, cuerpos lúteos, corpus albicans, folículos atrésicos y folículos fibrosos.
- La médula: es la zona más interna del ovario, rodeada por la corteza. Los límites entre corteza y médula no están muy bien definidos. La médula está formada por tejido conectivo fibroelástico laxo y contiene numerosos vasos sanguíneos, vasos linfáticos y nervios, los cuales entran y salen del ovario por una zona llamada hilio. En la médula no hay folículos ováricos. En ocasiones, existen ciertas formaciones tales como la rete ovarii procedente de invaginaciones primitivas del epitelio germinal.
La histología del ovario varía a lo largo del ciclo menstrual de la mujer madura. En la menopausia no se observan formaciones foliculares y el estroma contiene masas hialinas y fibrosas (corpus albicans y folículos atrésicos).
b. Irrigación e inervación del ovario
- Arterias. Las arterias del ovario proceden de la arteria ovárica y de la arteria uterina. La arteria ovárica, al alcanzar el ovario, da origen a una rama tubárica lateral que aborda la trompa uterina. La arteria ovárica desciende de inmediato por el hilio del ovario, donde da origen a algunas ramas que penetran en el ovario. Cuando llega a la extremidad uterina de este órgano, se anastomosa con la arteria uterina, dando origen a nuevas ramas ováricas.
- Venas. Forman en el hilio y en el mesoovario un plexo muy desarrollado. Las ramas que nacen de este plexo se dirigen a las venas ováricas y uterinas.
- Vasos linfáticos. Siguen el trayecto de los vasos ováricos, y drenan a la derecha en los nódulos linfáticos aórticos laterales subyacentes al pedículo renal, y a la izquierda en los nódulos linfáticos pregóticos y aórticos laterales subyacentes al pedículo renal y vecinos a este. Además, un colector linfático drena habitualmente, tanto a la derecha como a la izquierda, en un nódulo linfático medio de los nódulos linfáticos ilíacos externos.
- Nervios. Proceden del plexo intermesentérico por medio del plexo ovárico, que acompaña a la arteria ovárica. Los nervios destinados al ovario constituyen un plexo periarterial denso, en el que los filetes nerviosos se anastomosan en todos los sentidos y penetran en su trayecto pequeñas masas ganglionares. Esto son nervios vasomotores, pero también nervios sensitivos, que transmiten las sensaciones dolorosas nacidas en el ovario.
Trompas de Falopio
Las trompas ponen en comunicación la cavidad uterina con la cavidad abdominal e indirectamente con el ovario.
a. Anatomía de las trompas
Tienen un aspecto más o menos cilíndrico con una longitud de 10 a 14 cm, un diámetro externo de 1-1,5 cm y una luz del tubo que varía entre 1-4 mm de diámetro.
Desde el punto de vista anatómico está dividida en cuatro porciones:
- Porción intramural: esta porción está incluida, en su mayoría, dentro del músculo uterino y atraviesa el miometrio en 1 a 2 cm. Es la porción intrauterina más estrecha de la trompa y la cual se comunica con la cavidad del útero.
- Porción ístmica: de 2 a 4 cm y constituye la porción extrauterina más estrecha del oviducto.
- Porción ampular: de 4 a 6 cm de largo y su diámetro va aumentando progresivamente hasta el infundíbulo. Se considera la porción más ancha y larga de la trompa.
- Infundíbulo: cerca del ovario y lo rodea con unas proyecciones digitales, en forma de mano, que abraza al ovario en el momento de la ovulación y son las llamadas “fimbrias”.
b. Constitución de las trompas
Están constituidas por tres capas:
- La externa o peritoneo que constituye el meso de la trompa y posteriormente el ligamento ancho.
- La capa media o muscular está constituida por fibras musculares lisas, que se disponen a su vez en dos capas, la interna y la externa.
- La capa interna o mucosa. La mayor parte de las trompas, a excepción de la porción intramural de las trompas, están contenidas en los ligamentos anchos del útero y ocupa el borde superior de estos ligamentos (mesosálpinx).
c. Irrigación e inervación de las trompas de Falopio:
- Arterias. Las arterias de la trompa uterina proceden de dos orígenes: de
la uterina y de la ovárica. Estas arterias se anastomosan en el ligamento
ancho siguiendo la línea de soldadura de mesosálpinx y el mesoovario o
aleta posterior.
- Anastomosis no desdobladas: es posible describir tres arterias tubáricas: una arteria tubárica interna o ítsmica; una arteria tubárica externa o ampollar; una arteria tubárica media o intermedia.
- Anastomosis desdobladas: cuando la anastomosis de la ovárica o de la uterina es doble, cada uno de los dos arcos anastomóticos marchan aisladamente. El arco anterior está en el mesosálpinx, en el que suministra las tubáricas interna y media.
- Venas. Las venas nacidas de las redes capilares de las dos túnicas, muscular y mucosa, se dirigen hacia el mesosálpinx y forman, por su anastomosis, una red de mallas muy anchas, alargadas paralelamente al eje de la trompa. Finalmente, van a desembocar a las venas uteroováricas. Los vasos linfáticos drenan en los ganglios lumbares.
- Nervios. Provienen del plexo ovárico y del plexo uterino.
C. Útero
El útero es un órgano intrapélvico situado en el centro de la excavación pelviana entre la vejiga y el recto, cuya misión más importante es albergar el huevo fecundado durante toda la gestación.
El útero es un órgano hueco. Sus dimensiones varían con la edad y con la paridad de la mujer, pero por término medio oscila entre 6 y 9 cm, su anchura entre 3 y 4 cm y su diámetro anteroposterior entre 2 y 3 cm. El peso del útero no grávido oscila entre los 70-100 g y ocupa el espacio de la pelvis entre la vejiga y el ano.
El útero se fija a las paredes de la pelvis a través de tres pares de ligamentos: los ligamentos laterales o ligamentos anchos del útero, los ligamentos anteriores o ligamentos redondos del útero y los ligamentos posteriores o pliegues rectouterinos. Los ligamentos anchos del útero se dividen en dos partes: una superior, denominada mesosálpinx y, otra inferior, que constituye el mesometrio o base del ligamento ancho del útero. Tanto los ligamentos redondos como los pliegues rectouterinos pueden considerarse expansiones de los ligamentos anchos del útero.
a. Constitución del útero
Se compone de dos porciones anatómicofuncionales distintas, el cuerpo y el cuello, están separadas por una zona, el istmo, que fuera del embarazo no presenta relevancia anatómica.
- Cuerpo uterino: ocupa la porción superior del útero y es una formación eminentemente muscular. En el centro posee una cavidad; en su extremidad superior tiene dos estrechamientos llamados cuernos del útero que se continúan con el istmo de las trompas, y en su extremidad inferior, a través del istmo, con el cuello uterino.
- Cuello uterino: tiene forma cilíndrica. Su longitud varía entre 2,5 y 3 cm. En su extremidad superior se continúa, insensiblemente con el istmo. Su extremidad inferior, cónica, termina protuyendo en la porción superior de la vagina.
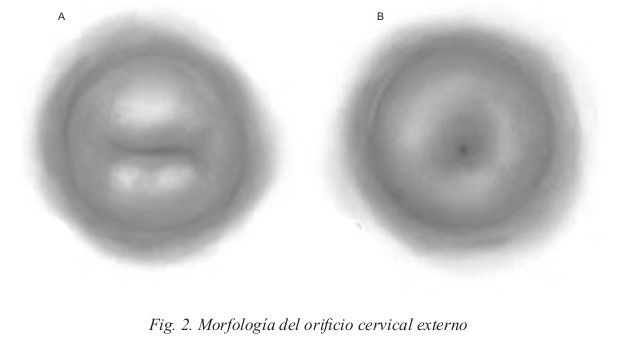
El cuerpo y el cuello uterino están tunelizados en su centro (cavidad cervical). Esta cavidad tiene forma cilíndrica y comunica la cavidad corpocervical con la vagina. La extremidad superior de la cavidad cervical o zona de unión con el istmo se denomina Orificio Cervical Interno OCI, y la extremidad inferior Orificio Cervical Externo OCE, que tiene morfología distinta según la paridad de la mujer: en las nulíparas es puntiforme (B) y en multíparas tiene una forma de hendidura transversal (A). Fig. 2.
b. Histología del cuerpo uterino
Está formado por tres capas bien diferenciadas:
- Peritoneo: recubre el cuerpo uterino en sus caras anterior y posterior en toda su extensión. Adhiriéndose fuertemente a él, resultando difícil la separación del miometrio.
- Miometrio: es la capa más voluminosa. Está constituida por fibras musculares lisas y tejido conjuntivo.
- Endometrio: capa mucosa que recubre el cuerpo uterino, esta mucosa responde con cambios morfológicos importantes a los estímulos hormonales del ovario y posee la curiosa propiedad de descamarse periódicamente cada 28 días, aunque la zona necrosada se regenera rápidamente.
c. Histología del cuello
Está constituido fundamentalmente por tejido conjuntivo fibroso. Contiene fibras musculares lisas en proporción muy inferior al cuerpo uterino. Estos haces musculares se localizan sobre todo en zonas más periféricas del cuello.
El cuello uterino está revestido en su superficie por dos tipos de epitelios totalmente distintos, cuya constitución es de gran relevancia ya que plantea problemas importantes en la práctica diaria ginecológica.
La zona del cuello uterino que se pone en contacto con la vagina denominada ectocérvix o portio, está revestida por epitelio escamoso estratificado, mientras que la cavidad endocervical o endocérvix, está revestido por un epitelio cilíndrico con numerosas glándulas.
d. Irrigación e inervación del útero
- Arterias. El útero recibe sus arterias de la arteria uterina. Un poco antes de llegar al cuello, la arteria uterina proporciona: ramas vesicovaginales (para vejiga y vagina) y una arteria cervicovaginal destinada a la parte inferior del cuello y a la pared anterolateral de la vagina.
- Venas. Las venas uterinas vierten en los densos plexos uterinos situados a los lados del útero. Estos plexos, anastomosados superiormente con las venas ováricas, vierten en los troncos ilíacos internos por medio de las venas uterinas.
- Vasos linfáticos. Se deben distinguir entre los vasos linfáticos del útero y los del cuello. Los del cuello del útero se dirigen mayormente en los nódulos ilíacos externos intermedios y superiores, y a veces a un nódulo linfático ilíaco común del promontorio. Los vasos linfáticos del cuerpo uterino, drenan en los nódulos linfáticos aórticos laterales, en los nódulos linfáticos preaórticos próximos al origen de la arteria mesentérica inferior, en los nódulos linfáticos ilíacos externos intermedios y, a veces, en los nódulos linfáticos inguinales superiores, superficiales y profundos, por medio de un vaso satélite del ligamento redondo del útero.
- Inervación. El útero está inervado por el plexo uterino que nace del borde anterior del plexo hipogástrico inferior.
Vagina
La vagina es un conducto que pone en comunicación la cavidad uterina con la vulva.
Tiene una longitud que oscila entre 8-12 cm. Podemos distinguirle dos caras: anterior y posterior; dos caras laterales o bordes y dos extremidades, superior e inferior. Si se observa al corte transversal, ambas caras vaginales se encuentran en contacto, lo que demuestra que la vagina es un conducto aplanado de delante a atrás.
a. Constitución de la vagina
La pared vaginal está constituida por tres capas:
- La capa externa, formada por la fascia vaginal, a través de la cual la vagina se relaciona por delante con la uretra y con la porción inferior de la vejiga urinaria, y por detrás con el recto y el peritoneo que recubre la porción más inferior de la cavidad pelviana.
- La capa media está constituida por fibras musculares lisas que se disponen a su vez en dos capas: una interna con fibras circulares y otra externa con fibras longitudinales.
- La capa interna o mucosa, está constituida por un epitelio plano estratificado.
b. Irrigación e inervación de la vagina
- Arterias. En la parte superior y media del conducto vaginal se encuentran ramas vaginales que derivan de la arteria uterina. La parte inferior del conducto vaginal está irrigada por ramas de la arteria rectal media.
- Venas. Forman, a ambos lados de la vagina, el plexo venoso vaginal. Estas venas, en parte, vierten directamente en la vena ilíaca interna o bien en uno de sus afluentes.
- Vasos linfáticos. Pueden seguir dos caminos, o bien drenan en los nódulos linfáticos ilíacos externos o bien en nódulos linfáticos ilíacos internos y comunes del promontorio.
- Nervios. Proceden del plexo hipogástrico inferior.
Órganos genitales externos
Se designa así al conjunto de órganos ubicados debajo de la pared abdominal anterior, en el perineo anterior, por delante del ano, por dentro y arriba de la cara medial de los muslos. Coronada por el monte del pubis (de Venus), la vulva, las formaciones labiales, el aparato eréctil y las glándulas anexas.

A. Monte de Venus
Es un saliente redondeado, situado debajo de la pared abdominal, delante de la sínfisis pubiana, en la parte anterior de la vulva. Se cubre de pelos en la pubertad. Está formada por un conjunto de tejido celulograsoso donde terminan las fibras de los ligamentos redondos del útero. Su sensibilidad depende de las ramas genitales del plexo lumbar, iliohipogástrico e ilioinguinal.
B. Formaciones labiales
Son los labios mayores y menores, en número de cuatro, dos de cada lado.
- Labios mayores: es un pliegue cutáneo, alargado de adelante hacia atrás, en el cual se reconoce:
- Una cara lateral: muy pigmentada, con pelos y separada del muslo por el surco genitocrural.
- Una cara medial: situada en contacto con el labio opuesto y separada del labio menor homólogo por el surco interlabial.
Los labios mayores se unen por sus extremos; su unión posterior constituye la horquilla o comisura posterior, situada inmediatamente por delante de la fosa del vestíbulo. El labio mayor está cubierto por un tegumento delgado bajo el cual se encuentra una capa muscular lisa; el dartos labial, al que cubre un tejido celulograsoso más o menos abundante. La piel de la cara medial contiene glándulas sebáceas.
- Labios menores: es un pliegue cutaneomucoso situado medialmente al labio mayor, adelante y lateral a la hendidura vulvar. Su extremidad superior, antes de llegar al clítoris, se divide en dos hojas secundarias, anterior y posterior; la cara posterior, corta, se dirige a la cara posterior del clítoris donde se inserta formando con la opuesta el frenillo del clítoris, la anterior, más larga, pasa delante del clítoris y se reúne en la línea mediana con el pliegue similar opuesto formando al órgano eréctil, una especie de envoltura semicilíndrica, el prepucio del clítoris. El revestimiento cutaneomucoso posee un armazón fibroelástico y contiene numerosas glándulas sebáceas.
C. Espacio interlabial
Aparece cuando se separan los labios. Por debajo del clítoris comprende de adelante hacia atrás: el vestíbulo, el ostio externo de la uretra y el ostio vaginal.
D. Himen
En la mujer virgen constituye una especie de membrana cuya concavidad se dirige hacia el centro del ostio inferior de la vagina, estrechándolo en parte. Es un septo incompleto que limita los conductos vaginales y vulvar. De forma variable, pero en general podemos encontrar tres tipos: semilunar, anular y labiado.
E. Aparato eréctil
Comprende el clítoris y los bulbos vestibulares.
- Clítoris: es el homólogo del pene en el hombre; aparece como un saliente submucoso, arriba del vestíbulo. Sólo es visible la extremidad anterior, única y mediana de los cuerpos cavernosos adosados entre sí. Las envolturas del clítoris están formadas por la piel, tapizada por una capa fibroelástica. Los cuerpos cavernosos están constituidos por un tejido eréctil de grandes mallas vasculares; en cambio el glande no es sino conjuntivo, con revestimiento mucocutáneo de extrema sensibilidad.
- Bulbos vestibulares: son formaciones eréctiles bilaterales, en contacto con la fascia inferior del diafragma urogenital; ocupan el borde adherente de los labios menores y cuyo borde interno limita con los ostios vaginal y uretral.
F. Glándulas anexas
a. Glándulas anexas uretrales y periuretrales
Son poco desarrolladas.
b. Glándulas de Bartolino
Son glándulas situadas a cada lado, en la parte posterolateral del ostio vaginal. Están medialmente debajo de la mucosa; lateralmente en relación con el bulbo vestibular. Su conducto excretor se abre en la base de los labios menores. Estas glándulas, que se desarrollan en la pubertad, segregan un líquido que lubrica las partes genitales durante el coito.
c. Glándulas de Skene
Son dos glándulas que cumplen la misma función que las anteriores y se encuentran una a cada lado del orificio de la uretra.
Mamas
La glándula mamaria constituye la característica fundamental de los mamíferos, quienes alimentan a sus crías con el producto de su secreción, la leche. En casi todos los mamíferos la vida del recién nacido depende de la capacidad de su madre para amamantarlo, por lo tanto, una lactancia adecuada es esencial para completar el proceso de la reproducción y la supervivencia de la especie. En general, la leche de una especie no permite asegurar la supervivencia de las crías de otra especie.
La histología de la glándula mamaria es prácticamente la misma en todas las especies: un parénquima glandular, compuesto de alvéolos y ductos, y un estroma de soporte. Cada célula alveolar se comporta como una unidad de secreción, produciendo leche completa, sintetizando y transportando desde el plasma sanguíneo las proteínas, grasas, hidratos de carbono, sales minerales, anticuerpos y el agua, que son los principales constituyentes de la leche. El proceso de síntesis y de secreción es similar en todas las especies. La composición química de la leche y la disposición anatómica del sistema de almacenamiento de la leche en la glándula mamaria varía en las diversas especies.
A. Anatomía de la mama
Las glándulas mamarias están presentes en ambos sexos. En el hombre se mantienen rudimentarias toda la vida, en cambio en la mujer están poco desarrolladas hasta antes de la pubertad, cuando empieza el proceso de maduración. El máximo desarrollo de estas glándulas se produce durante el embarazo y especialmente en el período posterior al parto, durante la lactancia.
La mayor parte de la masa de la mama está constituida por tejido glandular y adiposo. Durante el embarazo y la lactancia el tamaño de la mama aumenta debido al crecimiento del tejido glandular.
La base de la glándula mamaria se extiende, en la mayoría de los casos, desde la segunda hasta la sexta costilla, desde el borde externo del esternón hasta la línea axilar media. El área externo de cada glándula se extiende hacia la axila y se denomina “prolongación axilar”. La cara profunda de la mama es ligeramente cóncava y se encuentra en relación con el músculo pectoral mayor, el serrato anterior y la parte superior del oblicuo externo del abdomen. La mama está separada de estos músculos por la aponeurosis profunda. Entre ambas, hay un tejido areolar laxo denominado espacio retromamario; éste permite que la mama tenga cierta movilidad sobre la aponeurosis profunda que cubre al plano muscular.
La cara superficial de la mama está cubierta por piel. Aproximadamente en el centro de esta cara se encuentra el pezón que está en relación al cuarto espacio intercostal en la nulípara. La base del pezón está rodeada por una zona de piel hiperpigmentada, de 2.5 cm denominada areola. El pezón contiene numerosas fibras musculares lisas, en su mayoría de tipo circular, las que se contraen al estimularlo mecánicamente, originando la erección del pezón. La areola posee numerosas glándulas sebáceas, entre ellas es posible reconocer algunas que durante el embarazo y la lactancia determinan levantamientos de la piel de la areola, denominadas glándulas de Montgomery; éstas contienen estructuras histológicas similares a la parte glandular de la mama y producen una secreción grasa que lubrica el pezón y la areola. Bajo la areola se ubican las dilataciones de los conductos galactóforos llamadas senos lactíferos, que acumulan leche y el niño debe exprimir al mamar.
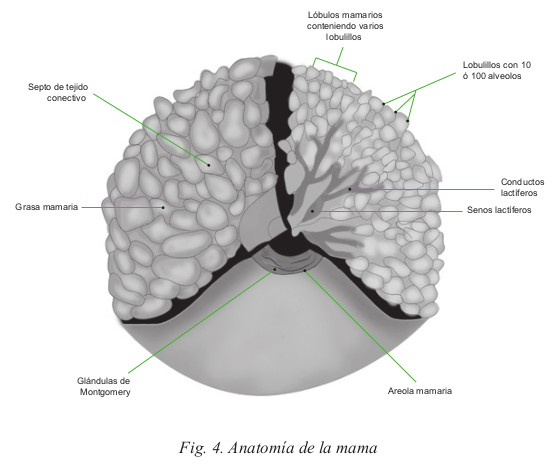
B. Estructura de la glándula mamaria
La glándula mamaria está formada por tres tipos de tejidos: glandular de tipo tubuloalveolar, conjuntivo que conecta los lóbulos, y adiposo que ocupa los espacios interlobulares. El tejido celular subcutáneo rodea la glándula sin que exista una cápsula claramente definida, desde éste se dirigen hacia el interior numerosos tabiques de tejido conectivo. Estos tabiques constituyen los ligamentos suspensorios de la mama o ligamentos de Cooper.
Un conjunto de quince a veinte lóbulos mamarios conforman la glándula mamaria, cada uno con su aparato excretor, que se abre en el pezón por medio de un conducto lactífero. Los lóbulos mamarios están constituidos por numerosos lobulillos que se encuentran unidos entre sí por tejido conectivo, vasos sanguíneos y por su sistema excretor, los conductos lactíferos (Fig. 5).
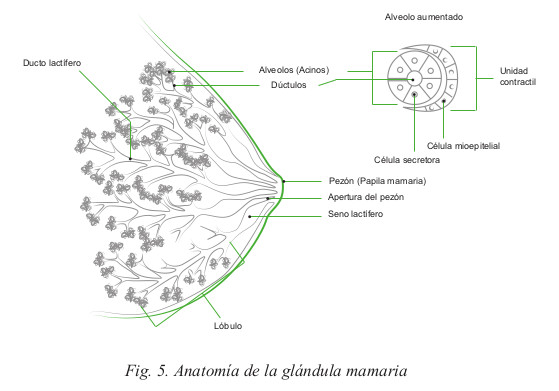
Los lobulillos están formados por diez a cien acinos, cada cual con su conducto excretor denominado conducto terminal. Los acinos están estructurados por un conjunto de células secretoras que producen la secreción láctea y conforman una cavidad a la cual vierten esta secreción; están rodeados de células mioepiteliales y capilares sanguíneos de singular importancia en el proceso de secreción y eyección de la leche.
El sistema de conductos lactíferos que vacía la glándula mamaria es el siguiente: el acino se vacía a través de un conducto terminal, el cual converge con sus congéneres para formar el conducto lobulillar, que recoge la secreción láctea de todos los acinos de un lobulillo. Los conductos lobulillares se reúnen para formar el conducto interlobulillar, que al unirse con otros conductos de este tipo, forma el conducto lobular o segmentario, de mayor calibre que los anteriores, que se dirige al pezón y antes de llegar a él, bajo la areola mamaria, se dilata formando el seno lactífero, el que se angosta nuevamente al desembocar en el pezón.
Los conductos están revestidos por epitelio cuboideo o cilíndrico. Por fuera de este epitelio, entre él y la membrana basal, existe una capa de células mioepiteliales muy ramificadas, que envuelven a los conductos y acinos. En los conductos de mayor tamaño, el epitelio consta de dos o más capas de células, que cerca del orificio externo del pezón, se transforman en epitelio plano estratificado.
La estructura de la glándula mamaria varía con la edad y es influenciada por el embarazo y la lactancia. Antes de la pubertad, la mama posee unos pocos conductos rudimentarios cubiertos en su interior epitelio plano y envuelto en tejido conectivo. Después de la pubertad, debido a la influencia de las hormonas ováricas, especialmente los estrógenos, los conductos se comienzan a ramificar y en sus extremos se forman pequeñas masas sólidas, esféricas, de células poliédricas, que constituirán los alveolos. Durante el estado de reposo, el epitelio glandular está separado del estroma vascularizado vecino por una fina zona de fibroblastos, a través de los cuales no penetran vasos. Esta unión epitelio-estromal, posiblemente, ejerce un control sobre el paso de sustancias a las células secretoras. Los alveolos activos sólo aparecen durante el embarazo, período en el cual, los conductos se ramifican y en su parte terminal se forma un lumen que aumenta de tamaño a medida que se va cargando de secreción.
Simultáneamente, aumenta la cantidad de tejido adiposo y la irrigación de la mama. En las últimas semanas del embarazo, la secreción adquiere características especiales y se denomina calostro. Algunos días después del parto, aparece la verdadera secreción láctea, la que distiende los alveolos que en ese momento están tapizados por una sola capa de células cilíndricas bajas. A medida que aumenta la cantidad de secreción, las células se aplanan, desapareciendo los espacios intercelulares o desmosomas. Durante el período de secreción el citoplasma de las células es basófilo, al microscopio electrónico se observan mitocondrias, lisosomas y ribosomas libres. Encima del núcleo, que se sitúa en la parte más basal de la célula, está el aparato de Golgi al que acompañan grandes vacuolas proteicas y lipídicas.
Después de la menopausia la glándula mamaria se atrofia y los elementos celulares de los alveolos y conductos degeneran, y disminuyen en número.
C. Irrigación de la mama
a. Arterias
Las arterias responsables de la irrigación de la mama son las siguientes:
- Mamaria interna: esta arteria se origina de la subclavia, desciende por la cara posterior de los cartílagos costales hasta el sexto o séptimo espacio intercostal. En su trayecto, la mamaria interna origina ramas intercostales anteriores, que transcurren por los espacios intercostales, uniéndose con las intercostales posteriores que vienen directamente de la aorta. Son las ramas intercostales anteriores las que producen múltiples vasos perforantes que pasan por los espacios intercostales y penetran en la glándula mamaria, desde la profundidad y de adentro hacia afuera. Son responsables aproximadamente del 50 % de la irrigación arterial, nutriendo el músculo pectoral, los cuadrantes internos, la piel y el complejo areola- pezón, alrededor del cual constituyen una red periareolar.
- Torácica lateral: también denominada torácica inferior, nace hacia el final de la subclavia aún por detrás del pectoral menor o bien directamente de la arteria axilar. Con un trayecto descendente, da múltiples ramas que atraviesan el pectoral al cual irrigan, hasta penetrar la glándula mamaria. La torácica lateral es denominada por algunos autores, arteria mamaria externa.
- Arterias intercostales posteriores: estas arterias son ramas directas de la aorta, transcurren desde atrás a lo largo de los espacios intercostales, emitiendo numerosas ramas perforantes para irrigar la pared del tórax. Algunas ramas penetran el pectoral y el serrato y llegan a la glándula mamaria.
- Arteria acromio-torácica: es una rama de la axilar, que da una colateral hacia la articulación acromio-clavicular y otras descendente hacia la región interpectoral. Transcurre entre ambos músculos dando varias ramas hacia la cara posterior de la mama.
- Arteria subescapular y torácica superior: se trata de otras ramas de la subclavia y axilar, que en su trayecto emite múltiples ramas hacia la cara anterior del tórax.
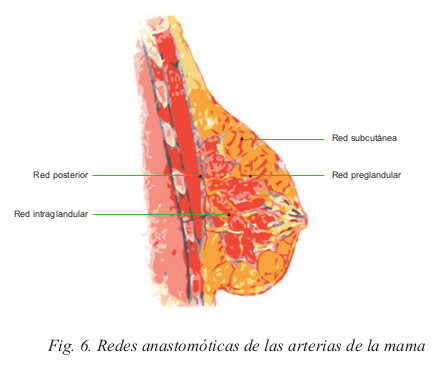
El mayor aporte sanguíneo está representado por la mamaria interna, la torácica lateral y las intercostales posteriores. La distribución de las ramas de todas estas arterias se produce en forma de 2 pedículos principales, llamados pedículo supero externo y supero interno, y 3 pedículos accesorios, llamados ínfero externo, ínfero interno y superior.
Estos 5 pedículos mantienen numerosas anastomosis que constituyen 4 redes diferentes:
- Red subcutánea
- Red preglandular
- Red intraglandular y
- Red retroglandular (Fig. 6)
Las arteriales se distribuyen en forma radial, desde la periferia hacia el centro de la mama, por lo que se considera que las incisiones radiales afectan menos la circulación. Sin embargo, en la práctica, las incisiones arciformes se realizan sin ninguna limitación, incluso a nivel periareolar donde existe un círculo anastomótico que permite incisiones hasta del 50 % de la circunferencia areolar. Desde el punto de vista práctico, es importante comprender que la rica red de anastomosis arteriales permite cualquier tipo de intervención e incisión cutánea sin riesgo de necrosis.
Esta rica red vascular se ramifica ampliamente hasta formar los delgados capilares que rodean los acinos mamarios, donde se produce el retorno sanguíneo y comienzan a formarse las venas mamarias.
b. Drenaje venoso de la mama
El retorno venoso de la glándula mamaria, presenta características muy particulares, ya que su distribución no siempre es paralela a la irrigación arterial. Existe una red venosa superficial que se inicia por debajo del pezón y la areola, localización donde recibe el nombre de plexo venoso de Haller. De este plexo nacen numerosas ramas que, en todas direcciones, discurren por el tejido subcutáneo hasta desembocar en la vena mamaria interna y torácica lateral (mamaria externa). Algunas ramas discurren hacia arriba y desembocan en las venas superficiales del cuello. Los troncos venosos de la mamaria interna y la torácica inferior, van paralelos a las arterias y llegan a la vena axilar y subclavia respectivamente.
Por el contrario, la red venosa profunda presenta una distribución similar a la irrigación arterial; la mayor parte de estas venas se dirigen en dirección a la pared torácica, penetran los espacios intercostales para drenar hacia las venas intercostales y la mamaria interna. Desde aquí, la sangre procedente de la mama pasa a la subclavia o bien hacia las venas vertebrales y de allí a la ácigos.
Las metástasis hematógenas en el cáncer de mama son poco frecuentes; sin embargo, es bien conocido que las costillas, vértebras e incluso los huesos de la pelvis pueden presentar metástasis distantes, y es el drenaje venoso, a través de las intercostales y la vena ácigos, la vía preferente para este patrón de diseminación.
c. Drenaje linfático de la mama
La diseminación de la enfermedad maligna de la mama se produce preferentemente a través de los vasos linfáticos, de aquí que resulte especialmente importante el conocimiento de la distribución de la red linfática y la ubicación de los ganglios que drenan la mama. Los vasos linfáticos de la mama son subcutáneos o glandulares. Los más numerosos son aquellos que drenan los cuadrantes externos, y se dirigen fundamentalmente hacia la axila. Los que drenan los cuadrantes internos se dirigen hacia la vía de la mamaria interna, mientras que los de la areola y el pezón, drenan hacia ambos lados. Así, se pueden diferenciar dos vías principales de drenaje linfático: la vía axilar que claramente reviste mayor importancia y la vía mamaria interna.
Los estudios con radioisótopos han logrado identificar claramente que el 75 % del drenaje linfático de la mama sigue la vía axilar, mientras que cerca del 22 % sigue la vía mamaria interna. El 3 % restante se dirige directamente a través de las vías intercostal, supraclavicular o hacia la mama contralateral.
La confluencia de numerosos vasos linfáticos de la mama en dirección a la axila, produce de 2 a 3 troncos principales que atraviesan la fascia de la base de la axila, a partir de la cual se distribuyen en forma progresiva a lo largo de todos los ganglios axilares.
Rouviere propone que, el drenaje linfático de la glándula mamaria se produce en forma centrípeta, siguiendo un patrón según el cual desde la profundidad del parénquima los vasos linfáticos se dirigen hacia la superficie donde forman el llamado plexo linfático subareolar de Sapper, del cual se forman los troncos que llevan la linfa en dirección a la axila o la mamaria interna.
Sin embargo, Haagensen plantea que el patrón de drenaje linfático es centrífugo, y a partir de la glándula mamaria, se produce la confluencia de los linfáticos menores hasta formar los troncos principales ya descritos. Para Haagensen, la existencia del plexo de Sapper no significa que recoja todo el drenaje de la mama, sino que es responsable del drenaje de un área especialmente importante como lo es el complejo areola-pezón y parte de la glándula subyacente.
d. Inervación de la mama
La sensibilidad superficial de la mama es responsabilidad de los 6 primeros nervios intercostales, y de la rama supraclavicular del plexo braquial. Al igual que la irrigación arterial y el drenaje venoso y linfático, en el complejo areola-pezón existe una importante red nerviosa que confiere especial sensibilidad a esta zona y es responsable de la erección del pezón ante los estímulos sexuales y la succión, así como del reflejo de la lactogénesis y la lactopoyesis.
Órganos pélvicos relacionados
En la pelvis, además de los órganos reproductores internos, la mujer, posee órganos pertenecientes al aparato urinario (los uréteres, la vejiga y la uretra) y al aparato digestivo (recto y ano). Éstos son los órganos pélvicos relacionados con los órganos genitales (Fig. 7). Los órganos pélvicos se mantienen en su sitio debido a los músculos del piso pélvico que son un conjunto de músculos y ligamentos que cierran la cavidad abdominal en su parte inferior. Varias capas de tejido conectivo que se llaman fascias también los apoyan.
A. Vejiga urinaria
Tiene forma esférica cuando está llena y de tetraedro cuando está vacía. Está situada por detrás de la sínfisis del pubis y por delante del útero, de la vagina y del recto. El vértice anteroinferior corresponde al orificio uretral. En los vértices superoexternos desembocan los uréteres. Su superficie superior está tapizada por el peritoneo, en contacto con el cuerpo y cuello uterino, formando así el fondo de saco anterior o vesicouterino. El espacio de Retzius se sitúa entre el pubis y la vejiga urinaria, cubierto por tejido adiposo extraperitoneal.
La irrigación arterial proviene de la arteria ilíaca y de sus ramas: arteria umbilical en la parte superior, la arteria vesical inferior en su parte media y la arteria pudenda en su parte inferior.
La drenaje venoso se lleva a cabo por el plexo venoso pélvico que recubre el espacio pre-vesical en su parte posteroinferior y termina en la vena hipogástrica.
La inervación está constituida por el plexo hipogástrico (simpático) y por el plexo pre-sacro (parasimpático).
B. Uréteres
Son dos tubos musculares de unos 25-30 cm de longitud. Comunican los riñones con la vejiga. Son dos estructuras retroperitoneales cuya mitad inferior se encuentran en la pelvis. El uréter penetra en la pelvis cruzando por delante de la bifurcación de la arteria ilíaca común y alcanza el suelo pélvico siguiendo la pared lateral de la pelvis. A nivel de la espina isquiática se dirige hacia delante y hacia dentro, por debajo del ligamento ancho y entre las arterias uterinas y vaginales, para alcanzar el fórnix vaginal lateral. En este punto se encuentra a 2 cm del cérvix. Después asciende por delante de la vagina hasta alcanzar el trígono vesical, penetrando en la vejiga en sentido oblicuo.
La irrigación arterial la constituye las arterias ilíacas y las arterias uterinas.
El drenaje venoso se corresponde con las venas ureterales que acompañan a las arterias y reciben los mismos nombres.
La inervación se lleva a cabo gracias a los nervios ureterales derivados de los plexos nerviosos adyacentes: rectal, aórtico e hipogástrico. Los nervios espinales son los L1-L2.
C. Uretra
Conducto muscular de unos 4 cm, en la mujer, por donde pasa la orina desde la vejiga al exterior. Sigue un curso antero-inferior desde el orificio uretral interno. En su parte inferior, se dirige a la sínfisis púbica. En la parte anterior desemboca en la vagina. La mitad inferior se encuentra en el perineo.
La irrigación arterial se corresponde con la arteria pudenda interna y la arteria vaginal.
El drenaje venoso se lleva a cabo por la vena pudenda y la vena vaginal. Éstas acompañan a las arterias.
La inervación se corresponde con el nervio pudendo.
D. Recto
Porción final del tubo digestivo. Por su parte superior, se continúa con el colon sigmoides a la altura de la S-3. Por debajo, con el conducto anal para terminar en el ano. Tiene tres curvas: superior, inferior y media (hacia la izquierda).
La irrigación arterial se lleva a cabo con las arterias rectales (superior e inferior).
El drenaje venoso se consigue gracias a las venas rectales (superior, media e inferior).
La inervación del recto y del ano es compleja y difiere de la inervación del resto del tubo digestivo. El ano-recto, está inervado por nervios sensitivos motores y otros dependientes del parasimpático como así también por el sistema nervioso entérico.
El nervio principal es el nervio pudendo que se origina del 2º, 3º y 4º nervio sacro e inerva al esfínter anal externo, a la mucosa anal y a la pared anorrectal.
E. Perineo
El perineo se halla limitado hacia delante por el arco púbico, lateralmente por las ramas isquiopubianas y las tuberosidades isquiáticas y hacia atrás por los ligamentos sacrociáticos mayor y menor. Todo ello, hace que tenga una forma romboidal. La línea transversa que une las tuberosidades isquiáticas, divide el perineo en un triángulo anterior urogenital, donde se encuentra el diafragma urogenital, y un triángulo posterior o anorrectal donde están las fosas isquiorrectales.
a. Perineo anterior
El perineo anterior está formado por tres planos musculoaponeuróticos: un plano profundo, un plano medio y un plano superficial:
- El plano profundo o diafragma pélvico, está constituido por una formación músculo tendinosa con forma de embudo, que sirve de sujeción a las vísceras pélvicas y abdominales. Incluye el músculo elevador del ano y el músculo coccígeo, recubiertos por una capa superior y otra inferior de fascia. El músculo elevador del ano se fija a la pelvis en las ramas descendentes y horizontales del pubis, espina ciática, aponeurosis del obturador interno y bordes del coxis. Sus fibras se dirigen a la línea media y se entrecruzan con las del lado opuesto. El músculo coccígeo se inserta en la espina ciática y sus ramas se abren en abanico fijándose en las caras laterales de los segmentos sacros inferiores y cóccix.
- El plano músculo aponeurótico medio, también llamado diafragma urogenital, se encuentra atravesado por la uretra y la vagina. Está formado por una capa superior y otra inferior que recubren los músculos perineales profundos, que se originan en la rama isquiopubiana y terminan en el triángulo rectovaginal, el músculo esfínter externo de la uretra y los vasos y nervios pudendos.
- El plano músculo aponeurótico superficial, incluye el músculo esfínter anal externo, los músculos isquiocavernosos, que se originan en la tuberosidad isquiática y terminan insertándose en las caras superior y lateral del clítoris; los músculos perineales transversos superficiales que se extienden desde la cara interna de la tuberosidad isquiática hasta el rafe, que va del ano a la comisura vulvar y, los músculos bulbocavernosos, que rodean el orificio inferior de la vagina y la terminación de la uretra.
b. El perineo posterior
El triángulo anal contiene el conducto anal y sus esfínteres, el cuerpo ano coccígeo y las fosas isquiorrectales, que contienen una gran cantidad de tejido adiposo, los vasos y nervios hemorroidales inferiores y los vasos y nervios pudendos internos.

La pelvis ósea
La pelvis es una estructura ósea situada entre la columna vertebral a la que soporta y las extremidades inferiores sobre las que se apoya. Este órgano óseo está constituido por la reunión de cuatro huesos: el sacro, por detrás y los dos coxales, formados primitivamente por el ilion por arriba y fuera, el isquion por abajo y el pubis por delante (Fig. 8). Estos huesos se relacionan entre sí por cuatro articulaciones:
- Articulación sacroilíaca: (derecha e izquierda): es una sínfisis estática o de muy poca movilidad.
- Articulación interpúbica: Es una sínfisis. Dispone de un manguito fibroso periarticular y fibrocartílago interóseo. Su relajación en el embarazo le proporciona una extensibilidad de 3-4 mm necesarios para el parto. Tras el parto regresa a su estado normal.
- Articulación sacrococcígea: Es del tipo anfiartrosis y permite movimientos de flexión y extensión. Dispone de un disco interarticular y un fuerte sistema ligamentario (anterior, posterior y lateral) cuyo reblandecimiento durante la gestación facilita el aumento del diámetro anteroposterior en 2-3 cm a este nivel, en el momento del parto.
A. Partes de la pelvis
La pelvis se divide en dos partes: por un lado está la pelvis falsa, superior o grande y, por otro lado, la pelvis verdadera, inferior o pequeña que constituye el canal óseo del parto, estando ocupado por recto, vejiga y los órganos reproductores en la mujer.
La pelvis verdadera se encuentra delimitada por:
El sacro, el cual está compuesto, normalmente, de cinco vértebras fusionadas (a veces, como en los tipos pélvicos antropoides presentan seis vértebras). Morfológicamente, es una pirámide con la base orientada hacia delante y arriba, el vértice hacia abajo y atrás y presentando, habitualmente, una concavidad interna o anterior. Su borde anterosuperior corresponde al cuerpo vertebral de la S-1 y se denomina promontorio.
La sínfisis púbica está constituida por el cartílago que une las dos ramas anteriores del pubis. La forma de esta articulación es un ovoide, donde el eje mayor es oblicuo hacia abajo y atrás formando un ángulo de 50° con la vertical. La sínfisis constituye la pared anterior de la pelvis menor.
a. El estrecho superior
El estrecho superior constituye el área que delimita superiormente la pelvis ósea verdadera u obstétrica.
Su perímetro es una línea que parte desde el promontorio por atrás, pasa por los alerones del sacro, las líneas innominadas y termina en la sínfisis púbica por delante.
El concepto de este término hace referencia a un verdadero canal, ya que no todas las estructuras óseas expuestas están al mismo nivel.
Los diámetros del estrecho superior son:
- Diámetro antero-posterior: es la distancia desde al promontorio al pubis.
Dentro de este diámetro hay diferentes referencias diametrales según los
puntos que se tomen como referencia.
- Conjugado verdadero: se mide desde la parte de la articulación púbica más interna o próxima al promontorio.
- Conjugado obstétrico: constituye el conjugado real ya que es la distancia más corta entre las estructuras óseas antes citadas.
- Conjugado diagonal: mide hasta el borde inferior del pubis.
- Segundo conjugado: distancia entre el punto más interno del pubis y la articulación S-1/S-2.
- Diámetro transverso: es la mayor distancia entre dos puntos de la línea innominada y perpendicular al diámetro anteroposterior. Su tamaño y posición varía según la forma del estrecho superior.
- Diámetros oblicuos: son dos derecho o izquierdo. Se extienden desde la articulación sacroilíaca hasta la tuberosidad iliopubiana contralateral.
b. El estrecho inferior
El estrecho inferior está limitado posteriormente por sacro y cóccix y lateralmente por las espinas ciáticas.
Tiene forma romboidal. Sus lados posteriores están definidos por los ligamentos sacrociáticos, mayor y menor, y el músculo isquiococcígeo. Sus lados anteriores son las ramas isquiopubianas unidas en la línea media por el ligamento púbico inferior. Se describen el diámetro anteroposterior (que separa el vértice del cóccix del punto medio de la sínfisis del pubis) y el diámetro transversal que separa las tuberosidades isquiáticas. Es el diámetro menor de la pelvis. De esta manera tiene una trascendental importancia durante el parto.
La excavación pélvica, es el hueco de la pelvis verdadera, delimitada en su parte superior por el estrecho superior, en la inferior por el estrecho inferior, en la anterior por el pubis, en la posterior por el sacro y lateralmente por los huesos coxales.

c. Osteometría pélvica
Estos conceptos son la base de la osteometría pélvica. Existen otros parámetros que son:
- Plano mayor: espacio comprendido entre el estrecho superior y el estrecho medio.
- Plano menor: coincide con el estrecho inferior.
- Sagital posterior: diámetro anteroposterior del estrecho superior situado por detrás de la intersección con el diámetro transverso obstétrico.
Las líneas perpendiculares que imaginariamente se levantan en el punto medio de cada plano de la pelvis se conocen como ejes. Así, tenemos:
- El eje del estrecho superior está inclinado hacia abajo y atrás, y su prolongación pasaría por el ombligo y por el cóccix.
- El eje del estrecho inferior si se prolongara incidiría en el promontorio sacro.
De esta forma, se puede calcular el eje en cualquier plano intermedio, y la resultante de todos ellos formaría el eje pélvico. Este eje pélvico es el camino que debería seguir el feto durante el parto, y constituiría el canal del parto propiamente dicho.
Todas las pelvis presentan una inclinación hacia delante, formando respecto de la horizontal, un ángulo de 60-70º el estrecho superior y un ángulo de 15º el estrecho inferior, estando por ello las partes posteriores de cada plano más elevadas que las anteriores.
B. Planos pélvicos
Nos permite seguir la altura de la presentación fetal a lo largo del canal del parto. Es como un sistema de coordenadas, en los que se dan puntos en las tres dimensiones del espacio. Son cuatro, todos paralelos:
- Primero: coincide con el estrecho superior.
- Segundo: pasa por el punto inferior de la sínfisis del pubis y a la altura de la S-2/S-3.
- Tercero: a la altura de las espinas ciáticas.
- Cuarto: coincide con el extremo caudal del cóccix.
C. Tipos de pelvis
Los criterios seguidos para su clasificación se basan en la morfología del estrecho superior. Trazando el diámetro transverso se divide éste en dos áreas: el segmento posterior (que determina el tipo de pelvis) y el anterior (que determina la tendencia) describiéndose académicamente cuatro tipos de pelvis (Fig. 9):
a. Pelvis ginecoide
Se caracteriza por la posición del diámetro transverso centrada, de tal manera que el sagital posterior es similar al anterior, lo que le confiere una forma ovalada o redondeada. Las paredes laterales son verticales y las espinas ciáticas no sobresalen demasiado. El arco púbico es ancho. Es sin duda el tipo de pelvis más frecuente en el sexo femenino. Esta morfología es la más fisiológica para el parto.
b. Pelvis androide
El diámetro transverso está retrasado por lo que el sagital posterior es más corto que el anterior. El sacro está adelantado en la pelvis y el ángulo púbico es agudo (similar a un corazón de naipes francés invertido). Las paredes son convergentes y las espinas ciáticas prominentes. Es el más pésimo de los tipos pélvicos para facilitar el parto.
c. Pelvis antropoide
Tiene forma de óvalo con su eje mayor en sentido anteroposterior. Las paredes laterales presentan convergencia aunque no muy pronunciada. Suele asociarse a un sacro recto y con seis vértebras lo que confiere una gran profundidad a este tipo de pelvis. Las espinas isquiáticas suelen ser prominentes y el arco púbico algo estrecho.
d. Pelvis platipeloide
Se caracteriza por tener un diámetro transverso, aunque centrado, muy ancho en relación al anteroposterior lo que le confiere una morfología de pelvis aplanada. El sacro es excavado y corto y el arco púbico es muy ancho. Es la más rara de las pelvis.

Fisiología del aparato reproductor femenino.
Ciclo reproductor femenino: ciclo ovárico y ciclo endometrial
La capacidad de reproducción es, desde el punto de vista biológico, la función específica propia de la mujer. Esta capacidad aparece en la pubertad, cuando se desarrollan las condiciones anatómicas y fisiológicas necesarias para el establecimiento del ciclo sexual femenino. El ciclo ovárico y endometrial son dos fenómenos simultáneos, que en circunstancias normales afectan cíclicamente al ovario y al endometrio de la mujer no embarazada durante su periodo reproductivo. La finalidad del ciclo ovárico es proporcionar un óvulo para la fecundación. La finalidad del ciclo endometrial es suministrar un emplazamiento adecuado donde el óvulo fecundado pueda implantarse y desarrollarse con el fin primordial de obtener un nuevo ser.
A. Ciclo ovárico
El ovario debe considerarse como una estructura dinámica con dos funciones íntimamente relacionadas:
- Gametogénica: con producción de ovocitos haploides fecundables.
- Endocrina: con producción de hormonas esteroideas con el fin de crear las modificaciones adecuadas para la nidación y nutrición del huevo o cigoto.
Estas dos funciones son sinérgicas y, aún considerando la función de la secreción esteroidea, es necesario tener en cuenta que la misión primordial del ovario es la producción de óvulos fecundables, estando todas las actividades ováricas subordinadas a ella. Por lo tanto, el ovario es el regulador del ciclo genital femenino, ya que mediante las variaciones de su producción estrogénica modula la liberación hipofisaria de gonadotropinas, y determina su patrón cíclico de secreción. No obstante, para un adecuado desarrollo del ciclo, es necesaria una estrecha relación funcional entre el hipotálamo, la hipófisis y el ovario. Si los niveles de gonadotropinas son bajos se interrumpe la foliculogénesis y, si son excesivamente altos se produce una superovulación, al alterarse los mecanismos ováricos de selección folicular.
a. Foliculogénesis
El elemento fundamental del ciclo ovárico es el folículo, el cual permanece en el ovario desde las primeras semanas de desarrollo embrionario. Las oogonias (células endodérmicas originadas en el saco vitelino), migran hasta la cresta genital en la quinta semana de gestación. Allí, comienzan un proceso de mitosis, multiplicándose considerablemente, y otro de meiosis, lo que permite la aparición de los primeros folículos primordiales en la semana dieciséis de gestación. Los folículos primordiales están constituidos por un ovocito.
En el momento del nacimiento, el ovario tiene entre uno y dos millones de folículos primordiales, que en la pubertad se han reducido a trescientos mil, y sólo unos cuatrocientos llegarán a la ovulación.
Habitualmente, la fase folicular tiene como resultado el desarrollo y la permanencia de un folículo maduro. El proceso se completa en 10-14 días. Durante éste se dan lugar a una serie de fenómenos hormonales y de péptidos sobre el folículo, que hace que a partir de un folículo primordial obtengamos un folículo maduro, habiendo éste pasado por las etapas de preantral, antral y preovulatorio.
Es desconocido el mecanismo por el cual se decide cuántos y cuáles folículos comienzan a crecer en un ciclo. Del grupo seleccionado sólo habrá un ganador.
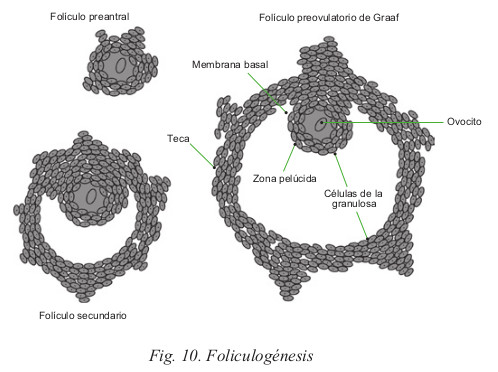
El folículo primordial es el estado inicial, el cual está constituido por una célula central o célula germinal rodeada por una capa continua de células en una sola hilera. El número de folículos primordiales varía a lo largo de la vida de la mujer; al nacer, el ovario contiene por término medio de 250.000 a 500.000, estos folículos van disminuyendo con la edad, entre los 40 y 44 años habrá aproximadamente 8.300. Sin embargo, sólo algunos cientos, entorno a unos 400, alcanzan la madurez definitiva, se transforma en folículo de Graaf, y posteriormente, tras la liberación del ovocito, en cuerpo amarillo.
Al iniciarse un nuevo ciclo, varios folículos primordiales comienzan a madurar aunque habitualmente sólo uno alcanzará la madurez total. Al crecer el folículo primordial, la capa granulosa que rodea al ovocito prolifera y se dispone en varias hileras. Ya, en el folículo primario, aparecen receptores de FSH, estradiol y andrógenos. Posteriormente, aparece en el centro del folículo una cavidad, que desplaza la célula germinal a un extremo. Por fuera de la capa granulosa, se distingue una capa conjuntiva llamada teca, donde se distinguen dos zonas:
- Una interna, con abundantes vasos
- Otra externa, o teca externa
La aparición de la teca vascular es de gran importancia en la foliculogénesis, ya que es entonces cuando el folículo puede responder a las hormonas proteicas de la sangre periférica. Esta dependencia es característica y diferencia el folículo primario del secundario.
Cuando el folículo alcanza la madurez definitiva, se denomina folículo preovulatorio o folículo de Graaf, donde se distinguen las siguientes formaciones: cavidad central llamada Antro y capa granulosa. En una zona del folículo estas células se proyectan hacia el interior del antro. En el interior de esta acumulación se encuentra el ovocito, este grupo de células se denomina disco oóforo.
La hilera de células granulosas que se ponen en contacto con el ovocito se llama corona radiada y entre ésta y las células germinales existe una zona denominada membrana pelúcida (Fig. 10).
b. Ovulación
Se produce hacia el día 14 del ciclo. Se caracteriza por la salida del ovocito, rodeado por algunas células granulosas y acompañado de líquido folicular. El folículo de Graaf crece hasta alcanzar un diámetro de 10 a 12 mm. Se aproxima de forma gradual a la superficie del ovario, hasta que sobresale de éste, de manera que por compresión se va formando una zona clara y delgada que acaba por romperse, expulsa el líquido folicular junto con el óvulo, rodeado por la zona pelúcida y la corona radiada. Todo este proceso es precedido de una serie de modificaciones vasculares en la zona de ruptura. En el centro de la zona, aparece una formación en cono, adelgazada, con posterior separación y necrosis de la capa germinativa.
La superficie del folículo se hernia en esta zona y termina por romperse, permitiendo la salida del contenido folicular, es decir, se produce la ovulación. El motivo desencadenante puede atribuirse al aumento de la secreción de gonadotropinas LH (Hormona Luteinizante) y FSH (Hormona Folículoestimulante). El aumento brusco de LH a las diez o doce horas desencadena la ovulación.
El brusco episodio de aumento de LH, condiciona la aparición de tres acontecimientos fundamentales:
- Estimulación de la reanudación de la meiosis y expulsión del primer corpúsculo polar, lo que completa la primera fase reduccional de la meiosis.
- Luteinización y secreción no solo de estrógenos, sino también de progesterona. Ésta última provoca también una disminución de LH.
- Contribución al mecanismo de ruptura folicular por aumento intrafolicular de prostaglandinas.
Otros factores que intervienen en el mecanismo de la ovulación son:
- Enzimas proteolíticas. Las células de la teca y la granulosa segregan cantidades importantes de activador del plasminógeno durante el periodo periovulatorio. La plasmina y, probablemente otras enzimas, como la colagenasa, provocan la digestión enzimática de la pared del folículo.
- Mucificación del complejo ovocito-cúmulo. Posiblemente, por la acción conjunta de la FSH y LH, el citado complejo se separa del resto de la granulosa y queda libre en el líquido folicular.
- Aumento de la presión intrafolicular.
c. Formación del cuerpo lúteo o cuerpo amarillo
Después de la ovulación, la pared folicular se retrae por disminución de la presión en su interior. La teca interna presenta abundantes hemorragias y la cavidad también se encuentra ocupada por sangre, el folículo se colapsa y a partir este momento, comienza la fase del cuerpo lúteo, en la que se distinguen cuatro estados:
Estado de proliferación
La capa granulosa está constituida por células con poco citoplasma y sin vasos; llama la atención el engrosamiento de la teca interna, formadas por células grandes con abundante citoplasma. La teca interna contiene numerosos vasos, en la luz del folículo colapsado no se observa sangre.
Estado de vascularización
Las células de la capa granulosa crecen y se transforman. Estas células constituyen las típicas células luteínicas, penetrando en ellas vasos procedentes de la teca interna. Por el contrario, las células de la teca interna disminuyen de volumen.
Estado de madurez
El cuerpo lúteo maduro destaca sobre la superficie del ovario, tiene una forma más o menos esférica con un diámetro de 1 a 2 cm y de color amarillento. Superficialmente, está constituida por una delgada capa de células tecales. Por dentro de ellas, una gruesa capa de células luteínicas. En el centro de esta formación, se observa habitualmente una capa de tejido laxo, pero en algunas ocasiones, puede observarse líquido hemorrágico.
Estado de regresión
Los signos morfológicos del comienzo de la regresión del cuerpo lúteo comienzan entre los días 23 y 26 del ciclo, aunque la secreción de esteroides empieza a disminuir ya a partir del día 22. Histológicamente, la regresión del cuerpo lúteo se manifiesta por fibrosis e hialinización de las células luteínicas; cuando se extiende a todo el cuerpo lúteo se forma el cuerpo albicans. Cuando el óvulo es fecundado, el cuerpo lúteo no involuciona y se transforma en cuerpo lúteo gravídico.

Los folículos primordiales, que entran en proliferación al principio del ciclo y que no evolucionan, empiezan la regresión mucho antes y forman folículos atrésicos, éstos experimentarán una involución semejante a la de cuerpo lúteo y formaran los cuerpos fibrosos.
B. Ciclo endometrial
El endometrio es la mucosa que recubre la cavidad uterina. Está constituido fundamentalmente por glándulas, vasos y estroma. Bajo la acción secuencial y combinada de los esteroides ováricos, estrógenos y progesterona, la mucosa endometrial experimenta una serie de cambios morfológicos cíclicos a lo largo del ciclo genital femenino y posee la propiedad de descamarse periódicamente cada 28 días y de regenerar rápidamente la zona descamada.
Morfológicamente el endometrio se puede dividir en dos capas:
- Capa basal: apenas sufre cambios cíclicos y es la encargada de la regeneración postmenstrual del endometrio. Está irrigado por ramas rectas de las arterias radiales.
- Capa funcional: es la que sufre los cambios cíclicos y se descama al final de cada ciclo si no ha existido fecundación. Está irrigada por los plexos vasculares que forman con las venas, las ramas espirales de las arterias radiales.
En el ciclo endometrial se distinguen las siguientes fases: fase proliferativa, fase secretora y fase de descamación (Fig. 12).
a. Fase de proliferación
La fase proliferativa está asociada al crecimiento folicular ovárico y al incremento de la secreción de estrógenos. Histológicamente, se observan cambios en cada una de las estructuras que constituyen la mucosa endometrial, conduciendo al final de esta fase a un endometrio hiperplásico secundario al hiperestrogenismo.
Esta fase se extiende desde el final de la menstruación (días 3-5 del ciclo) hasta la ovulación (días 13-15 del ciclo) y se produce la reconstrucción y crecimiento del endometrio a partir de la capa basal, que persiste sin descamarse después de la menstruación.
Al principio, el epitelio de superficie endometrial se regenera, haciendo cesar la hemorragia menstrual. La regeneración es completa al tercer o cuarto día del comienzo de la menstruación; se realiza a partir de los fondos de sacos glandulares que quedaron del ciclo anterior, revistiendo toda la superficie mucosa.
Posteriormente, se produce un crecimiento en espesor, que afecta a todos los componentes endometriales, glándulas, vasos y estroma. Las glándulas endometriales son entonces tubulares y rectilíneas. Al aumentar el espesor del endometrio, las glándulas aumentan su longitud y al final de la fase proliferativa, próximo al día 14 del ciclo, las glándulas son discretamente más largas que el espesor del endometrio. También se observa cierta dilatación de las luces glandulares.
El epitelio que reviste las glándulas es de tipo cilíndrico. Inmediatamente después de la regla es casi cúbico, con núcleos redondos situados en el centro de la célula. Entre los días 7 y 10 del ciclo comienzan a aparecer numerosas mitosis en las células epiteliales. Se observan abundantes células ciliadas que aumentan progresivamente durante la fase proliferativa, que alcanzan su máximo hacia la mitad del ciclo, para descender después en la fase secretora. El epitelio de superficie experimenta modificaciones semejantes a la del epitelio glandular.
El estroma aparece constituido por células que contienen escasos citoplasmas y núcleos fusiformes. A mediados de la fase de proliferación se observa, en determinadas ocasiones, un edema más o menos marcado. A partir del día 7 comienzan a observarse mitosis en las células del estroma.
b. Fase secretora
Después de la ovulación, el endometrio muestra una sensibilidad a la acción combinada de los estrógenos y la progesterona. El espesor del endometrio no se modifica a partir del máximo alcanzado en el periodo preovulatorio (5-6 mm), a pesar de que continúa la influencia estrogénica.
Histológicamente, es característico el cese de la proliferación endometrial, aproximadamente 3 días después de la ovulación.
Los cambios morfológicos son al principio más llamativos a nivel glandular, y en la fase secretora media y tardía, o premenstrual, las modificaciones se hacen más aparentes en el estroma.
Los primeros signos de respuesta a la progesterona segregada por el cuerpo lúteo aparecen hacia el día 17 del ciclo. Se forman unas pequeñas vacuolas en las células epiteliales que les confiere un aspecto como apolillado. Estas pequeñas vacuolas confluyen en una más grande subnuclear, que desplaza al núcleo hacia el polo apical. Alrededor del 18-19 día del ciclo, las vacuolas pasan del polo apical y el núcleo al polo basal, comienza a aparecer secreción en las luces glandulares, mientras el borde apical celular se hace irregular. Más tarde, las glándulas se dilatan por acúmulo de secreción y se hace cada vez más aparente el edema del estroma. Hacia el día 22 puede observarse la vascularización típica de esta fase secretora.
Las células del estroma se agrandan; su citoplasma se hace más abundante y sus límites celulares mal definidos, conteniendo glucógeno y lípidos abundantes. Esta transformación desigual del estroma, que comienza hacia el día 23, se hace más acusada alrededor de los vasos espirales. Mientras tanto, las glándulas se han hecho más tortuosas y dilatadas, con el típico aspecto de diente de sierra.
Hacia el final de la fase secretora, aparece un infiltrado leucocitario que se va haciendo cada vez más intenso, sin que ello signifique ningún proceso inflamatorio, sino un infiltrado fisiológico. Las glándulas están revestidas de un epitelio bajo y de aspecto deshilachado; el estroma está edematoso y manifiesta una intensa reacción residuoide y el infiltrado leucocitario. Los vasos forman una extensa red capilar por debajo del epitelio endometrial y también alrededor de las glándulas. Las venas presentan dilataciones y anastomosis arteriovenosas, con esfínteres precapilares que, para algunos autores, juegan un papel importante en el mecanismo de la menstruación.
c. Fase de descamación
Empiezan a aparecer pequeños hematomas y el endometrio comienza a descamarse, dando lugar a la menstruación.
Si el óvulo es fecundado, este endometrio se prepara para albergar al huevo fecundado; pero si el huevo no es fecundado, el endometrio se descama, sobreviniendo la menstruación.
Durante la menstruación el endometrio se desprende de la capa funcional quedando solo la capa basal a partir de la cual se produce la regeneración.

Sistema hormonal femenino
Existen tres jerarquías diferentes de hormonas (Fig. 13):
A. Hormonas de liberación hipotalámica GnRH (Hormona Reguladora de la secreción de las Gonadotropinas FSH y LH)
Esta hormona estimula la hipófisis anterior (adenohipófisis) y produce la liberación de forma pulsátil de FSH y LH, de tal forma que los pulsos lentos sobreestimulan FSH, y los rápidos sobreestimulan LH y, la liberación continua desensibiliza los receptores e inhibe por tanto la FSH como la LH. A su vez, el hipotálamo está controlado por el SNC; la dopamina inhibe la producción de GNRH.
B. Hormonas de la hipófisis anterior (u Hormonas gonadotrópicas)
Son hormonas estimuladas por la secreción pulsátil de la GNRH:
- FSH (Hormona Estimulante del Folículo). Estimulan el crecimiento de los folículos en la capa granulosa del ovario y la producción de estrógenos. La liberación de FSH tiene dos fases: una primera meseta, se libera en la primera mitad de la fase proliferativa, y tiene como misión el crecimiento de la cohorte folicular y la selección del folículo dominante. El segundo pico sucede justo antes de la ovulación y estimula el crecimiento de la capa granulosa en el folículo que ha seleccionado; induce la actividad aromatasa en la granulosa que convierte los andrógenos en estrógenos y aumenta los receptores de FSH en la granulosa. La FSH es inhibida por la inhibina folicular y los estrógenos. Es decir, los estrógenos producidos gracias a la FSH, inhiben a la propia FSH mediante un feedback negativo.
- LH (Hormona Luteinizante). Su liberación tiene un sólo pico; el pico ovulatorio, consecuencia del efecto gatillo de los estrógenos. La ovulación es la consecuencia del pico de LH. La LH estimula el crecimiento de la teca, que produce andrógenos y favorece la luteinización del folículo tras la ovulación.
C. Hormonas ováricas
Estrógenos y Progesterona son las hormonas secretadas por el ovario en reacción al estímulo de las hormonas de la hipófisis anterior. La progesterona es la hormona por excelencia del embarazo, se sintetiza en los ovarios, cuerpo amarillo y placenta; es una hormona enlentecedora de todos los fluidos corporales y sistemas, bilis, saliva, tubo digestivo, sistema renal-urinario.

Función de las hormonas femeninas
A. Hormonas ováricas
- Estrógenos. Se producen básicamente en el ovario desde la menarquia hasta la menopausia. También se producen, pero en menor cantidad, en las glándulas suprarrenales y en la grasa. Fomentan sobre todo la proliferación y el crecimiento celular de zonas específicas del cuerpo (piel, hueso, aparato genital) y se encargan de la mayor parte de los caracteres secundarios de la mujer. El estrógeno más importante es el estradiol E2. En el embarazo, la placenta se encarga de producir estrógenos.
- Progestágenos. Se producen también en el ovario y se relacionan fundamentalmente con la preparación del útero para la gestación y de las mamas para la lactancia. En el embarazo es también la placenta quien se encarga de su producción. El progestágeno más importante es la progesterona. Se pueden destacar otras funciones importantes de la progesterona que son la elevación de la temperatura corporal a partir de la ovulación (se utiliza en los estudios de fertilidad), disminución del moco cervical y en la gestación, disminución de la contractilidad uterina.
- Andrógenos. Hormona ovárica producida en el tejido estromal de las células de la teca (sufren aromatización y se transforman la mayoría en estrógenos).
B. Otras hormonas
Por último debemos hacer mención a otras hormonas importantes que se relacionan con el ciclo reproductivo de la mujer:
- Prolactina. Hormona hipofisaria, cuya acción fundamental es actuar sobre la mama para desencadenar la lactancia. Niveles altos de esta hormona producen anovulación, porque inhibe la GNRH.
- Oxitocina. Es una hormona que se almacena en la adenohipófisis o hipófisis anterior y su acción principal la realiza sobre el útero gestante produciendo la contracción de la fibra muscular lisa. Es importante en la lactancia porque produce la expulsión de la leche.
- HCG (Hormona Gonadotropina Coriónica Humana) Es una hormona placentaria. Tiene la función de mantener el cuerpo lúteo funcional, si existe gestación, hasta que la placenta adquiera la capacidad de producir estrógenos y progestágenos (7ª-10ª semana de gestación). Está presente en sangre y orina y su presencia diagnóstica la gestación (a partir 5ª semana en orina). Es una hormona exclusiva del embarazo.
- HPL (Lactógeno placentario). Es una hormona placentaria, cuya función principal es favorecer la lactogénesis y movilizar las grasas. Es una hormona exclusiva del embarazo. Es la antítesis de la insulina.
Causas de la menstruación
A. Causas hormonales
En el ciclo se producen la secreción de hormonas ováricas. Durante el desarrollo folicular se segregan estrógenos que inducen la fase de proliferación endometrial y, a partir de la ovulación, el cuerpo amarillo segrega estrógenos y progesterona, que conducen a la transformación secretora del endometrio.
Al final del ciclo ovárico, cuando el cuerpo amarillo envejece, se provoca una carencia hormonal de estrógenos y progesterona, pero fundamentalmente estrógenos, lo que condiciona la aparición de la menstruación.
B. Causas vasculares
Existen transformaciones vasculares que se producen al final de la fase secretora, donde se observa una gran complejidad vascular, con grandes dilataciones venosas a modo de senos venosos y anastomosis arteriovenosas. Se produce un espasmo del esfínter precapilar, dando lugar a una isquemia de las zonas irrigadas.
La sangre se deriva bruscamente a los vasos venosos dilatados y de paredes frágiles, que estallan. Como esto tiene lugar en zonas edematosas e isquémicas, se produce el desprendimiento de la mucosa, la menstruación.
C. Causas nerviosas
Las correlaciones hipotálamo-hipófisis-ovario y las del hipotálamo con la corteza hace que estímulos emocionales, interfieran en la regulación del ciclo y condiciona alteraciones del mismo, dando lugar a amenorreas o menstruación.
Variaciones y alteraciones del ciclo menstrual
El ciclo menstrual ideal tiene una periodicidad de 28 días, considerándose ciclos normales aquellos con intervalos mínimos de 24 días y máximo de 35 días. La duración del flujo menstrual más frecuente es de 3-6 días, aunque algunas mujeres normales tienen reglas de sólo 2 días de duración y otras de hasta 8 días. El volumen de sangre menstrual perdido es de unos 30 ml por ciclo, siendo anormal cuando supera los 80 ml.
A. Síndrome de tensión premenstrual
Se trata de un conjunto de síntomas que aparecen en las mujeres durante el periodo que precede a la regla, que afectan a su psiquismo, perturban su comportamiento y perjudican su actividad.
Entre los síntomas que pueden aparecer son: plenitud pélvica, sensación de peso en el abdomen, flatulencia, cefalea, irritabilidad, depresión, ansiedad, fatiga, agotamiento, náuseas, vómitos, diarreas y estreñimiento.
La causa de este síndrome no está clara pero principalmente se atribuye al desequilibrio hormonal que se produce en el ciclo sexual femenino.
El tratamiento consistiría en modificar hábitos alimenticios y de higiene y en la administración de progestágenos, antiprostaglandinas, y vitamina B-6, así como ACO (Anticonceptivos).
B. Dismenorrea
Se define como dolor pélvico o abdominal, inmediatamente antes de la menstruación o coincidiendo con ella. La etiopatogenia es desconocida, pero se asocia al aumento de la producción endometrial de prostaglandinas. Según su causa se clasifican en:
- Dismenorreas primarias aparecen 2 ó 3 años después de la menarquia y ocurre durante los primeros días del flujo. Se producen sin enfermedad orgánica demostrable. Son más frecuentes en nulíparas y su tratamiento se basa en administrar contraceptivos orales combinados, inhibidores de la síntesis de prostaglandinas ó analgésicos.
- Dismenorreas secundarias. Acompañan a alteraciones orgánicas del aparato reproductor de la mujer. Suelen aparecer después de comenzar la menstruación. Se asocia a endometriosis, enfermedad pélvica inflamatoria y anomalías anatómicas. El tratamiento sería referido a la patología de base que la produzca.
C. Amenorrea
Se define como la ausencia temporal o permanente de la menstruación. Se clasifica en amenorrea primaria y amenorrea secundaria.
- Amenorrea primaria: Cuando a los 18 años no se ha establecido la menstruación. Las causas más frecuentes son obstrucciones congénitas, ausencia de útero o alteraciones hormonales.
- Amenorrea secundaría. Cuando se interrumpe la menstruación ya establecida durante más de 3 meses. Existen amenorreas secundarias fisiológicas en el embarazo, lactancia y menopausia. Las causas más frecuentes son desequilibrios hormonales, alteraciones en nutrición, lesiones ováricas, enfermedades debilitantes, contraceptivos, administración de fármacos tranquilizantes, algunos síndromes como el de Cushing. El tratamiento en ambos casos depende de la causa.
D. Hemorragias uterinas anormales
Se definen como una alteración menstrual en exceso y se clasifican según aparecen en el tiempo en cíclicas y acíclicas y según su causa en orgánicas y disfuncionales.
Cíclicas:
- Hipermenorreas o menorragias. Se define como una menstruación con duración superior a la normal (más de 7 días) o en cantidad excesiva o ambas.
- Polimenorrea. Es una menstruación que aparece en ciclos cortos, con intervalos inferiores a 21 días, pero normal en cantidad y duración.
- Hiperpolimenorrea también llamada polimenorragia. Son menstruaciones con intervalos excesivamente frecuentes y cuya intensidad y duración son igualmente excesivas.
Acíclicas:
- Metrorragias: pérdida hemática de origen uterino, sin tener relación con el
ciclo. Pueden tener distinta intensidad:
- Spotting: pequeña pérdida hemática, tipo goteo, fuera de la menstruación.
- Orgánicas: existen patologías del aparato genital femenino que producen hemorragias (ver patología ginecológica).
- Disfuncionales. Son hemorragias anormales procedentes de la cavidad uterina, en ausencia de embarazo y no responden a ninguna causa orgánica. Se producen por alteraciones neuroendocrinas que afectan al ciclo sexual.
Otros términos relacionados con las alteraciones del ciclo menstrual son:
- Oligomenorrea que define a las menstruaciones de menos de dos días de duración.
- Ataxia menstrual. Con este término se hace referencia a la irregularidad menstrual.
Anatomía y fisiología de los órganos reproductores masculinos.
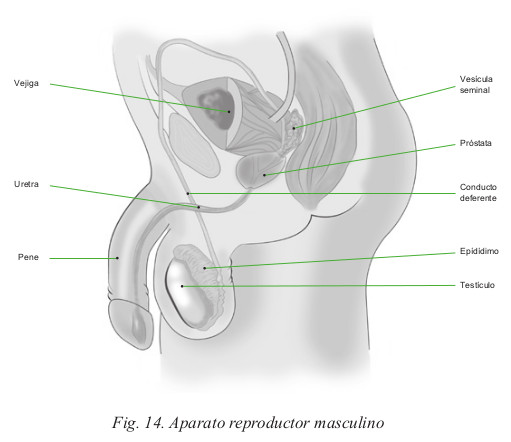
El aparato genital masculino está compuesto por los siguientes elementos (Fig.14):
- Los testículos.
- Los conductos genitales: epidídimo, conductos deferentes y conductos eyaculadores.
- La uretra.
- La próstata.
- Las vesículas seminales.
- El pene que contiene las estructuras eréctiles.
Testículos
Son los principales órganos reproductores del hombre. Producen los espermatozoides (Fig. 15)
A. Estructura, situación y función
- Los testículos son dos órganos situados en el exterior de la cavidad abdominal, el izquierdo un poco más bajo, debajo del pene y alojados en las bolsas escrotales o escroto. En su origen, en la vida embrionaria, los testículos se encuentran en el interior de la cavidad abdominal. La anormal permanencia de esta situación se denomina criptorquidia. Durante la fase embrionaria, los testículos migran desde el abdomen hacia su localización extracorporal a través del conducto inguinal. Al nacer, este trayecto queda sellado. Si no se cierra totalmente puede acumularse cierta cantidad de líquido, formando una bolsa quística que se denomina hidrocele.
- La forma de los testículos es oval, con un gran eje oblicuo por debajo y por detrás. Su longitud es de unos 5 cm y cada uno de ellos pesa alrededor de 20 g. La superficie del testículo es lisa y brillante, de color blanco, formada por una cubierta fibrosa denominada albugínea, muy tensa, lo que le confiere una consistencia dura.
- Cada testículo está subdividido en lobulillos, en un número de 200
a 300. Cada lobulillo contiene numerosos tubos muy delgados: los
túbulos seminíferos contorneados. Éstos albergan las células germinales
productoras de espermatozoides, las espermatogonias. El conjunto de los
túbulos termina en una red colectora denominada rete testis, desde donde
salen los conductillos eferentes, para desembocar en la primera porción
del conducto deferente, denominada epidídimo. En la pared de los túbulos
seminíferos contorneados se producen dos procesos esenciales:
- La espermatogénesis: proceso mediante el cual, a partir de una célula madre o espermatogonia con dotación diploide, aparecen cuatro células hijas o gametos masculinos con dotación haploide. Esta reducción del material genético es posible gracias a que en la gametogénesis se realiza una meiosis.
- La producción de testosterona: la principal hormona masculina: es sintetizada por células especializadas llamadas células de Leydig.
B. Tipos de células
Además de las células que participan directamente en la espermatogénesis, se observan otros dos tipos de células:
- Las células de Sertoli
- Las células de Leydig
Las células de Sertoli están situadas en la pared de los túbulos seminíferos. Estas células tienen un papel de soporte y de nutrición, y producen las sustancias químicas necesarias para la espermatogénesis.
Las células de Leydig se encuentran en el tejido conjuntivo que ocupan los espacios entre los túbulos seminíferos.
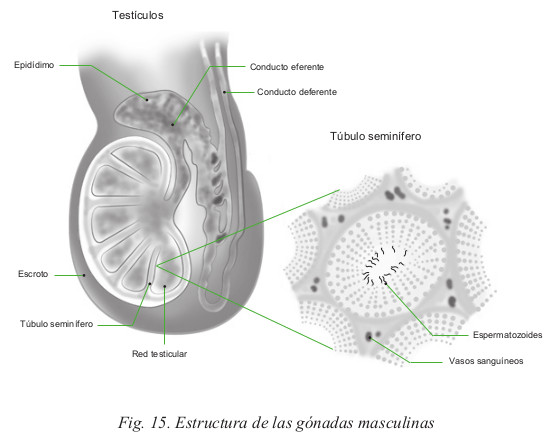
Escroto
Es la estructura de sostén de los testículos. Es un saco que consta de piel laxa y una fascia superficial que cuelga de la raíz o base del pene. Desde el exterior parece una sola bolsa de piel, separada en partes derecha e izquierda por un reborde mediano, el rafe, mientras que internamente el tabique escrotal lo divide en dos sacos, cada uno de los cuales contiene un testículo.
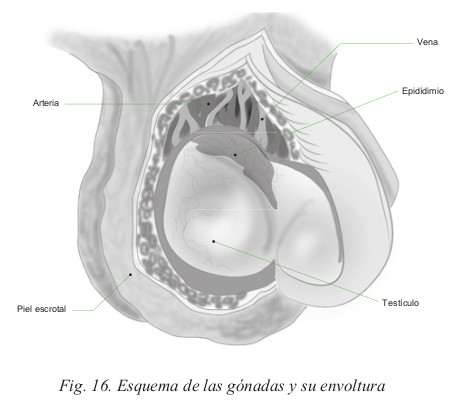
El tabique consiste en fascia superficial y tejido muscular, el llamado músculo dartos, que se compone de haces de fibra de músculo liso, también está presente en el tejido subcutáneo del escroto. Al contraerse, el dartos causa las arrugas en la piel del escroto. Se contrae en respuesta al frío y se relaja con el calor.
La localización del escroto y la contracción de sus fibras musculares regulan la temperatura de los testículos (2 ó 3 ºC menor que la temperatura corporal central), necesaria para la producción de espermatozoides. El músculo cremáster consiste en una pequeña banda de tejido muscular esquelético en el cordón espermático, eleva los testículos durante la exposición al frío y la excitación sexual, acercando los testículos a la cavidad pélvica, donde se invierte el proceso.
Los conductos genitales
A. Epidídimo
El epidídimo recubre el borde posterosuperior de cada testículo, tiene forma de signo de coma de unos 4-5 cm de longitud, aunque el conducto epididimario, que es una estructura muy contorneada, tiene en realidad unos 6 m de longitud al estirarlo.
a. Constitución
Está constituido por tres partes:
- Una cabeza: segmento dilatado situado en el polo superior donde los conductos eferentes del testículo se unen a los conductos del epidídimo.
- Un cuerpo: es la porción central angosta del epidídimo.
- Una cola: deshilada y más pequeña que se apoya en la parte inferior.
El conducto del epidídimo tiene un revestimiento de epitelio cilíndrico pseudoestratificado rodeado de capas de músculo liso.
Desde el punto de vista funcional: aumenta la motilidad de los espermatozoides a lo largo de 10 a 14 días. Los espermatozoides pueden permanecer almacenados en el conducto durante un mes o más.
B. Conductos deferentes
Existe un conducto deferente por cada uno de los testículos.
La cola del epidídimo, en su extremo más distal, se vuelve menos contorneada y aumenta su diámetro dando lugar al conducto deferente. El conjunto forma una curva llamada asa epidídimo-deferencial.
El conducto deferente es un canal largo de 45 a 50 cm de longitud, posee un revestimiento de epitelio cilíndrico pseudoestratificado y una cubierta de tres capas de músculo liso: la interna y externa son longitudinales y la intermedia circular.
El conducto deferente asciende hacia el polo superior del testículo a lo largo de la cara interna del epidídimo, sale de la bolsa escrotal hacia el conducto inguinal, y lo atraviesa penetrando en la cavidad abdominal. Circula sobre la cara lateral de la vejiga, pasa por encima del uréter y alcanza la base de la próstata, donde se dilata y se une a la desembocadura de las vesículas seminales. La porción dilatada del conducto deferente se llama ampolla.
Funciones: funcionalmente:
- Almacena espermatozoides, que conservan su viabilidad durante meses (los espermatozoides no eyaculados se reabsorben).
- Transporta células del epidídimo a la uretra mediante contracciones peristálticas de capa muscular.
C. Cordón espermático
Es una estructura de sostén del aparato reproductor masculino. Formado por el conjunto de vías que ascienden desde el testículo hasta el abdomen y que pasan por el conducto inguinal. Se distinguen:
- Las arterias espermáticas que provienen de la aorta.
- Las venas espermáticas: la vena espermática derecha va a parar a la vena cava inferior, la vena espermática izquierda desemboca en la vena renal izquierda.
- Los nervios vegetativos.
- El conducto deferente.
- Vasos linfáticos.
D. Conducto eyaculador
Los dos conductos eyaculadores tienen una longitud de unos 2 cm de longitud; se forman de la unión del conducto que proviene de la vesícula seminal y la ampolla del conducto deferente. Nace justo arriba de la base (porción superior) de la próstata, terminando en la porción prostática de la uretra donde vacía los espermatozoides y secreciones de las vesículas seminales antes de la eyaculación.
Vesículas seminales
Son dos glándulas en forma de saco de unos 5 cm, situadas en el plano posterior de la vejiga, en la base de ésta, por delante del recto. Las vesículas seminales derecha e izquierda reciben respectivamente los conductos deferentes derecho e izquierdo.
Secretan un líquido viscoso y alcalino. Su naturaleza alcalina ayuda a neutralizar el entorno ácido de la uretra masculina y del aparato reproductor femenino que inactivarían y matarían a los espermatozoides.
a. Composición del líquido
Este líquido contiene:
- Fructosa: la utilizan en la producción de ATP.
- Prostaglandinas: contribuyen a la motilidad y viabilidad de los espermatozoides.
- Proteínas de coagulación (diferentes de las sanguíneas).
El líquido que secretan comprende casi el 60 % del volumen del semen.
Uretra
En el hombre, la uretra es el conducto terminal compartido de los aparatos reproductor y urinario, ya que sirve para la expulsión tanto de semen como de orina. Su longitud es de unos 20 a 25 cm, cruza la próstata, el diafragma urogenital y el pene. Se divide en varios segmentos:
Partes de la uretra:
- El 1º segmento: segmento inicial que atraviesa la pared vesical.
- El 2º segmento: la uretra prostática, tiene de 2 a 3 cm de longitud y atraviesa la próstata.
- El 3er segmento: la uretra atraviesa el diafragma urogenital donde recibe el nombre de uretra membranosa. Ésta mide aproximadamente 1 cm.
- El 4º segmento: la uretra peneana o uretra esponjosa que cruza el cuerpo del pene, tiene unos 15-20 cm de longitud y termina en el meato urinario externo. En ella desembocan pequeñas glándulas mucosas a lo largo de todo el trayecto: las glándulas de Littre, con acción lubricante.
El par de glándulas de Cowper (bulbos uretrales), cada una del tamaño de un guisante, se sitúa en un plano inferior a la próstata, a ambos lados de la porción membranosa de la uretra dentro del diafragma urogenital y sus conductos se abren en la porción esponjosa. Estas glándulas secretan durante la excitación sexual una sustancia alcalina, que protege a los espermatozoides mediante la neutralización de ácidos de la orina en la uretra. También secretan moco, que lubrica el extremo del pene y el revestimiento de la uretra, esto hace que disminuya el número de espermatozoides que sufre daño durante la eyaculación.
Próstata
La próstata es una glándula que por su forma, tamaño, color y consistencia se asemeja a una castaña. En la edad adulta, su tamaño es de 3 cm de altura, 4 cm de anchura y 2 cm de grosor (puede incrementarse con la edad). Está situada debajo de la vejiga, rodeando la uretra y los conductos eyaculadores que desembocan en la uretra:
Orientación de la próstata:
- La base se orienta hacia arriba, bajo la vejiga.
- El vértice hacia abajo, se apoya en el diafragma urogenital (músculo transverso del perineo).
- Por detrás, está en relación con el recto (permite su exploración mediante el tacto rectal).
- Por delante está la sínfisis del pubis, de la que la separa la grasa y las venas prostáticas.
La próstata es una glándula de secreción exocrina que secreta un líquido blanquecino hacia la uretra, el liquido prostático, que se une a las secreciones de las vías espermáticas y los espermatozoides para constituir el semen. Este líquido es alcalino, por lo que neutraliza la acidez de los demás componentes del semen y aumenta la motilidad y fertilidad de los espermatozoides.
La próstata se contrae durante la eyaculación, junto con el conducto deferente y las vesículas seminales, expulsando su contenido a la uretra.
El semen
Es una mezcla de espermatozoides y líquido seminal; consiste a su vez en una mezcla de secreciones de los túbulos seminíferos, vesículas seminales, próstata y glándulas de Cowper.
El volumen de semen en cada eyaculación suele ser de 2.5 a 5 ml, con una concentración de espermatozoides de 50 a 150 millones por mililitro. Se requiere que los espermatozoides eyaculados sean tan numerosos porque solo una diminuta fracción llega al oocito secundario. Tiene un pH ligeramente alcalino (7.2 a 7.7). Las secreciones prostáticas le confieren un aspecto lechoso, mientras que las de vesículas seminales y glándulas de Cowper le dan al semen su consistencia adherente.
El semen contiene un antibiótico, la seminalplasmina, que mata ciertas bacterias.
El pene y las estructuras eréctiles
A. El pene
Órgano cilíndrico que pende sobre las bolsas escrotales. Es un conjunto de estructuras formado por los órganos eréctiles masculinos y las capas que lo rodean. Su tamaño y consistencia varían según se encuentre en estado de flacidez (unos 10 cm) o de erección (unos 15 cm).
Partes del pene:
- La raíz del pene: es la parte posterior, fijada al músculo transverso profundo del perineo y a las ramas isquiopúbicas.
- El cuerpo del pene: por debajo de la sínfisis púbica, la porción móvil del pene se despega.
- El glande del pene: es un abultamiento cónico que presenta el pene en su extremidad. Se abre a la uretra por un orificio llamado meato uretral. La base del glande es la corona, ésta se encuentra separada del cuerpo del pene por un surco. El prepucio es un repliegue cutáneo que rodea al glande y está fijado a éste en la cara inferior del pene por una brida cutánea, el frenillo del prepucio.
B. Cuerpos eréctiles
Tipos de cuerpos eréctiles. Se distinguen dos tipos de cuerpos eréctiles en el interior del pene:
- Los cuerpos cavernosos: son dos; éstos se fijan en las ramas isquiopubianas formando la raíz del pene. En este tramo están recubiertos por el músculo isquiocavernoso. Se unen bajo la sínfisis del pubis, desde donde emergen, y forman la porción dorsal del pene. Estos dos cuerpos se acercan uno al otro y se unen en una extremidad afilada recubierta por el glande.
- El cuerpo esponjoso: Se encuentra situado en la zona media del pene y mide alrededor de 15 cm; se fija bajo el músculo transverso profundo del perineo en un ensanchamiento denominado bulbo. Esta porción abultada, está rodeada por los dos músculos bulboesponjosos. A lo largo de su trayecto, el cuerpo esponjoso rodea la uretra; de esta forma, al salir de la próstata, la uretra desemboca en el bulbo del cuerpo esponjoso, y se convierte en la uretra peneana.
C. Capas del pene
Los cuerpos eréctiles del pene están rodeados por tres capas de tejido superpuestas.
Se distinguen desde dentro hacia afuera:
- La fascia peneana, es la capa más interna; es una envoltura fibroelástica que se continúa con la fascia superficial del escroto y perineo. Se une a la sínfisis del pubis por el ligamento suspensorio del pene.
- El músculo dartos del escroto se continúa también por el pene formando otra de sus envolturas. Es una capa de tejido muscular liso que recubre la fascia profunda del pene.
- La piel, con un tejido celular muy laxo; se adhiere al pene en toda su longitud, excepto en el glande, al cual se une por una línea en su cara inferior denominada frenillo. El resto de la piel del glande está libre, cubriéndolo solo en estado de flacidez. Esta porción de piel se denomina frenillo, durante la erección se retrae descubriendo el glande. Cuando su orificio anterior es cerrado y no permite la salida del glande, constituye la fimosis.

D. Función del pene
Tiene una doble función:
- Interviene en la micción, al contener en su interior parte de la uretra.
- Es el órgano copulador en el acto sexual.
Las hormonas del aparato reproductor masculino
A. La secreción hormonal del testículo
El testículo es una glándula mixta: exocrina y endocrina. Su función exocrina es la producción de espermatozoides. Su función endocrina es la secreción de la principal hormona masculina: la testosterona.
- Las células de Leydig, células especializadas del testículo, tienen como función secretar testosterona.
- El testículo fabrica otras hormonas androgénicas:
- La deshidroepiandrosterona (DHA)
- La androstenediona
De estas hormonas, la testosterona es la más activa e importante: origina el climaterio hormonal masculino. La testosterona depende de las hormonas hipotalámicas e hipofisarias.
El testículo también fabrica estrógenos a dosis reducidas.
B. Las funciones de la testosterona
Las principales funciones de la testosterona son:
- Sobre la espermatogénesis: actúa directamente sobre las células de Sertoli. Una deficiencia de testosterona causa esterilidad.
- Sobre el desarrollo del conjunto de los órganos genitales masculinos: testículos, epidídimo, conducto deferente, vesícula seminal, próstata y pene.
- Sobre los caracteres sexuales secundarios masculinos: desarrollo del sistema piloso, desarrollo de la masa muscular, aumento del timbre de la voz gracias al desarrollo de la laringe.
- Sobre el metabolismo: relacionada con los caracteres sexuales secundarios. Además de los órganos genitales y los senos, la morfología masculina es diferente de la morfología femenina. Esta diferencia es debida a un metabolismo diferente de los lípidos y de los prótidos.
C. La influencia hipotalamohipofisaria
El desarrollo y funcionamiento del testículo está bajo el control de hormonas hipofisarias, que a su vez están controladas por una hormona hipotalámica.
Las hormonas hipofisarias se llaman gonadoestimulinas, se distinguen:
- FSH: estimula el crecimiento de los túbulos seminíferos y la espermatogénesis por medio de las células de Sertoli.
- LH: estimula la secreción de testosterona actuando directamente sobre las células de Leydig.
La hormona hipotalámica implicada es la GnRH.
Tanto la hormona hipotalámica como las hipofisarias se mantienen en el hombre a niveles constantes.
Gametogénesis. Meiosis. Características del óvulo y del espermatozoide
Gametogénesis es el proceso inclusivo por el cual células diploides experimentan meiosis para producir gametos haploides altamente diferenciados y especializados.
Aunque la formación de gametos difiere en cada sexo, el espermatozoide y el óvulo son homólogos e involucran transformaciones morfofisiológicas.
La gametogénesis ocurre en las glándulas sexuales o gónadas, tanto en varón como mujer, cuando ellos han alcanzado la madurez sexual. Este momento conocido como pubertad, se caracteriza en los mamíferos por un desencadenamiento de una liberación basal continua de gonadotropinas (FSH y LH) en el hombre, o una liberación cíclica de ellas en la mujer. Estas hormonas actúan sobre sus órganos dianas, las gónadas, activando tanto la gametogénesis como la síntesis de esteroides gonadales. Estos esteroides actúan a su vez sobre las características sexuales tanto secundarias como accesorias, provocando los cambios típicos observados durante la pubertad.
La gametogénesis comprende 4 fases:
- Origen y migración de las gónadas
- Multiplicación de las células germinales
- Meiosis
- Maduración final antes de la fertilización
Meiosis
La meiosis es un mecanismo de división celular que permite la obtención de células haploides (n), con diferentes combinaciones de genes, a partir de células diploides (2n).
A. Objetivo biológico de la meiosis
La meiosis en el ser humano, está directamente relacionada con la sexualidad y tiene un profundo sentido para la supervivencia y evolución de las especies. El ser humano es considerado como la culminación de la evolución. De esta forma, la meiosis confiere a los organismos que la portan un par de características de indudable importancia que son:
- Reduce a la mitad el número de cromosomas: de esta manera, en los organismos con reproducción sexual, sobre todo en el reino animal, asegura que a cada nueva generación que aparece, el número de cromosomas sea constante, tras la fusión del gameto femenino con el masculino producida en la fecundación, impidiendo que haya alteración del número de cromosomas específico de cada ser vivo, característica que proporcionaría a las nuevas generaciones anomalías que podrían ser incompatibles con la vida, comprometiendo el futuro de la especie.
- Establece reestructuraciones en los cromosomas homólogos, mediante intercambios de material genético: esto supone una ventaja, ante hábitats cambiantes, ya que aumenta exponencialmente la variabilidad genética en la producción de gametos de un individuo, haciendo posible la aparición de combinaciones genéticas casi infinitas, permitiendo a las especies que poseen esta división celular aumentar su probabilidad de supervivencia.
B. Mecanismo de la meiosis
La meiosis (Fig. 18) consta de dos divisiones sucesivas de una célula diploide (2n) con una única replicación del ADN. El producto final son cuatro células haploides con n cromosomas. Así pues, consiste en una división del núcleo en dos secuencias, dando como resultado que el número diploide pase a ser haploide.
I División meiótica (reduccional)
A partir de una célula madre 2n se obtienen dos células hijas con n cromosomas. Se divide en varias fases:
Se le denomina también división reduccional, su duración representa el 90 % de toda la meiosis. En ella, los cromosomas homólogos se aparean e intercambian material genético entre ellos y posteriormente se separan reduciéndose el número de cromosomas a la mitad, de ahí el nombre. Se diferencian 4 etapas: profase I, metafase I, anafase I y telofase I.
Profase I
Es la etapa más larga, más compleja y más importante; en ella se diferencian 5 subetapas: leptoteno, zigoteno, paquiteno, diploteno y diacinesis.
- Leptoteno. Los cromosomas se condensan y se empiezan hacer visibles. Cada uno de ellos está formado por dos cromátidas estrechamente unidas, que no se distinguen hasta el final de la profase I. Cada cromosoma se une por sus extremos a la envoltura nuclear.
- Zigoteno. Los dos cromosomas homólogos de cada pareja se aparean longitudinalmente gen a gen; a este proceso de cromosomas homólogos apareados se les denomina bivalentes o tétradas (contiene 4 cromátidas).
- Paquiteno. En este período se produce el sobrecruzamiento o entrecruzamiento entre cromátidas homólogas, es decir, cromátidas no hermanas pertenecientes a la misma pareja de cromosomas homólogos. Mediante este proceso dos cromátidas homólogas se entrecruzan y posteriormente se rompen intercambiándose fragmentos entre ellas; como consecuencia se produce un intercambio de genes o recombinación genética, con ello aumenta la variabilidad.
- Diploteno. Los cromosomas homólogos comienzan a separarse, aunque permanecen unidos por unos puntos, llamados quiasmas, que se corresponden con los lugares donde se produjo el sobrecruzamiento.
- Diacinesis. En esta etapa se observan, por primera vez, las dos cromátidas que forman cada cromosoma que están unidas por el centrómero. Los pares de cromosomas homólogos permanecen unidos por los quiasmas que se establecen entre cromátidas homólogas.
Al final de este periodo desaparece l a sinapsis y se realiza mediante una estructura proteica denominada complejo sinaptinémico. A cada pareja a membrana nuclear y el nucleolo, y se empieza a formar el huso acromático. Los dos cinetocoros de cada cromosoma homólogo están fusionados y se sitúan en el mismo lado, a partir de ellos crecen los microtúbulos cinetocóricos.
Metafase I
El huso está totalmente formado. Las parejas de cromosomas homólogos (bivalentes) unidas por los quiasmas se sitúan en el ecuador del huso formando la placa metafásica.
Anafase I
Los quiasmas se rompen y los cromosomas homólogos de cada pareja comienzan a separarse, al ser arrastrados por las fibras cinetocóricas del huso que se acortan. Cada uno de estos cromosomas homólogos están formados por dos cromátidas y se dirigen hacia un polo de la célula; por consiguiente la mitad de los cromosomas irán a un polo y la otra mitad al otro.
Telofase I
Termina la migración de los cromosomas homólogos al polo correspondiente, y una vez allí sufren una cierta descondensación, se forma la membrana nuclear y el nucléolo, y desaparece el huso. Como resultado se habrán formado dos núcleos hijos que tendrán la mitad de cromosomas que el núcleo materno.
Inmediatamente se produce la citocinesis, obteniéndose dos células hijas que tendrán la mitad de cromosomas que tenía la célula madre, cada uno de estos cromosomas tendrá dos cromátidas.
II División meiótica (ecuacional)
Está precedida de una breve interfase, denominada intercinesis, en la que nunca hay duplicación del ADN. Es parecida a una división mitótica, constituida por la profase II, la metafase II, la anafase II y la telofase II con la única diferencia que se parte de células haploides (n) en vez de células diploides (2n).
Esta división se produce simultáneamente en las dos células hijas resultantes de la división anterior. Esta división es similar a una mitosis, en ella al igual que en la mitosis se separan las dos cromátidas hermanas de cada cromosoma. En esta división se diferencian cuatro etapas:
Profase II
Es muy breve, los cromosomas se condensan, desaparece la membrana nuclear, nucléolo y se forma el huso.
Metafase II
Los cromosomas tienen cada uno dos cromátidas unidas por el centrómero, se sitúan en el ecuador del huso formando la placa metafásica.
Anafase II
Se duplican los centrómeros y las dos cromátidas que forman cada cromosoma se separan, yendo cada una hacia un polo; cada una de ellas constituye un cromosoma hijo.
Telofase II
Termina la migración de los cromosomas, se descondensan, desaparece el huso y se forman las membranas nucleares, originándose dos núcleos. A continuación se divide el citoplasma. Como resultado se habrán formado 4 células hijas haploides (n) a partir de una célula diploide (2n). Estas cuatro células haploides serán genéticamente distintas entre sí ya que algunos de sus cromosomas están recombinados.
Ovogénesis
El desarrollo de los óvulos tiene lugar en las gónadas femeninas: los ovarios. En este órgano, las células madres germinales sufren un complicado proceso de división celular que se denomina ovogénesis, gracias a la meiosis.
Este proceso se inicia en la mayoría de las hembras de mamíferos durante la etapa embriofetal. Este temprano inicio determina muy precozmente el número definitivo de células germinales que irá desapareciendo hasta agotarse a medida que la mujer envejece.
La ovogénesis comienza poco después que las células germinales primordiales (CGP) colonizan la cresta genital. Estas células pierden su actividad migratoria. Luego empiezan a dividirse mitóticamente para dar origen a la ovogonias.
A. Fases de la ovogénesis
Fases de la ovogénesis relacionadas con la edad:
- Prenatal o período de proliferación. Empieza cuando las células germinales primordiales colonizan el lugar donde se originarán los ovarios; allí las células realizan divisiones mitóticas sucesivas y forman las ovogonias; éstas se encuentran en el interior de los folículos, al tercer mes de embarazo aumentan de tamaño y duplican sus cromosomas y originan los ovocitos de primer orden; se inicia la primera división meiótica.
- Postnatal o período de crecimiento. Comienza desde el nacimiento hasta el inicio de la pubertad. Los ovocitos de primer orden continúan en el mismo estado en que se detuvo la meiosis.
- Madurez sexual o periodo de maduración. Se inicia desde los comienzos de la pubertad hasta el momento en que la mujer experimenta los cambios debido a la menopausia. En la pubertad se caracteriza por la actividad del hipotálamo y la hipófisis; en este periodo se finaliza la meiosis de un ovocito de primer orden, produciéndose las divisiones sucesivas y continuas que caracterizan a esta división celular. El ovocito de primer orden (2n), entra en la división reduccional y se forman dos células hijas: ovocitos de segundo orden, célula haploide (n) y el corpúsculo polar de primer orden, célula haploide (n).
Resultado de la segunda división meiótica. Ambas células entran en la segunda división dando origen a:
- El ovocito de segundo orden, al óvulo (n) y un corpúsculo polar de segundo orden (n).
- El corpúsculo polar de primer orden a dos corpúsculos polares de segundo orden.
El resultado final de la ovogénesis es que se obtienen 4 células haploides (n), el óvulo y tres corpúsculos polares de segundo orden (n); éstos, los corpúsculos polares secundarios, no son funcionalmente aptos ya que carecen de los nutrientes necesarios y en consecuencia son absorbidos por el organismo femenino y no intervienen como células sexuales.
Proceso citológico de la ovogénesis. Sin tener en cuenta la diferencia de madurez sexual y considerando la ovogénesis como un proceso citológico de división celular sin ninguna pausa, las fases en las que se divide ésta son (Fig. 19):
- Fase de proliferación o multiplicación: las células madres germinales (2n) se multiplican por mitosis dando ovogonias (2n).
- Fase de crecimiento: las ovogonias atraviesan una fase de crecimiento y se convierten en ovocitos de primer orden (ovocitos I), también con 2n cromosomas. A diferencia de la espermatogénesis el crecimiento es considerable, ya que el óvulo es el gameto portador de la mayoría de las sustancias necesarias para el desarrollo del embrión.
- Fase de maduración: una vez que el ovocito primario ha completado su crecimiento, está ya preparado para atravesar las dos divisiones de la meiosis, y transformarse en una célula haploide con n cromosomas: la ovótida. Una peculiaridad muy importante de la ovogénesis, es que durante la meiosis el ovocito no se divide en cuatro células iguales, sino que la mayoría del citoplasma queda en una sola de ellas, la que dará lugar al óvulo. Así, cada ovocito primario da lugar a un único óvulo. Las otras tres células restantes, muy pequeñas, se denominan corpúsculos polares y se trata en realidad de gametos abortivos que permanecen un tiempo adosados al óvulo hasta que terminan por atrofiarse y desaparecer.
- Fase de diferenciación: la ovótida se transforma en el óvulo. En general no se trata de una fase de transformaciones tan acusadas como las que suceden en el espermatozoide. El óvulo es una célula haploide de gran tamaño, pues almacena sustancias nutritivas en forma de granos de vitelo. Como cualquier otra célula, está recubierto por la membrana plasmática. Pero además, existen otras membranas de gran espesor envolviendo a la membrana plasmática.
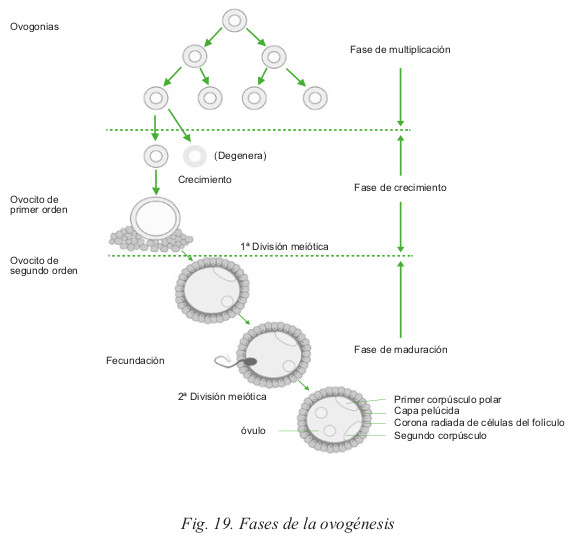
Espermatogénesis
La formación de los espermatozoides tiene lugar en las gónadas masculinas: los testículos. En el ser humano, los testículos son órganos compuestos por numerosos túbulos seminíferos que convergen en conductos comunes que llevan el esperma maduro al exterior. El examen microscópico de estos túbulos seminíferos permite reconocer fácilmente el curso de la espermatogénesis y distinguir sus diferentes fases (Fig. 20).
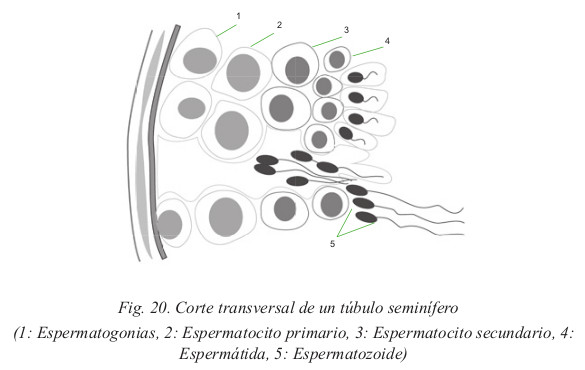
La espermatogénesis es la suma de los procesos que en el adulto terminan con la producción de espermatozoides, siendo este un proceso continuo que se inicia ya en la vida embrionaria. Se produce por la activación del eje hipotálamo-hipofisiario que involucra la secreción de la hormona estimulante del folículo (FSH) y la hormona luteinizante (LH).
A. Fases de la espermatogénesis relacionadas con la edad
La formación de los espermatozoides comienza en la pubertad y ocurre en el interior de los túbulos seminíferos mediante tres etapas:
- Prenatal o período proliferativo. En la tercera semana de gestación, se pueden reconocer las células primordiales germinales; éstas se dirigen hacia el lugar donde están los testículos, y allí sufren una división mitótica que da origen a los monocitos; éstos se multiplican y se originan las espermatogonias, que se ubican en la base de los túbulos seminíferos.
- Posnatal o período de crecimiento. Después del nacimiento, las espermatogonias fetales dan origen a las espermatogonias prepuberales o espermatocitos primarios, los cuales son células diploides (2n); éstas son las que originan los espermatozoides que se forman al pasar la pubertad.
- Madurez sexual o período de maduración: Cuando se activa el sistema hipotálamo-hipofisiario, se produce la formación de los espermatozoides; las espermatogonias mediante la mitosis, forman los espermatocitos de primer orden; estos pasan por un proceso de meiosis, antes de la primera división meiótica, los espermatocitos de primer orden aumentan de tamaño y sus cromosomas se duplican. Las células resultantes de la primera división meiótica, se llaman espermatocitos de segundo orden, luego ocurre la segunda división meiótica y se originan las espermátidas; éstas sufren modificaciones y producen los espermatozoides.
B. Proceso citológico de la espermatogénesis
Sin considerar, de igual manera que se ha hecho en la ovogénesis, la diferencia de madurez sexual y considerando la espermatogénesis como un proceso citológico de división celular sin ninguna pausa, las fases en las que se divide ésta son (Fig. 21):
- Fase de proliferación o multiplicación: pegadas a la pared del túbulo se encuentran unas pequeñas células (2n) que se multiplican activamente por mitosis, son las espermatogonias.
- Fase de crecimiento: las espermatogonias que quedan hacia la luz del túbulo experimentan una etapa de crecimiento y pasan a denominarse espermatocitos primarios o de primer orden (2n).
- Fase de maduración: los espermatocitos primarios van a sufrir la primera división de la meiosis, transformándose en espermatocitos secundarios (n). La segunda división de la meiosis produce unas células haploides llamadas espermátidas (n); por cada espermatocito primario se producen cuatro espermátidas.
- Fase de diferenciación o espermiogénesis: las espermátidas no son todavía los gametos; antes deben experimentar una serie de transformaciones anatómicas, etapa llamada espermiogénesis, al final de la cual quedarán convertidas en espermatozoides.
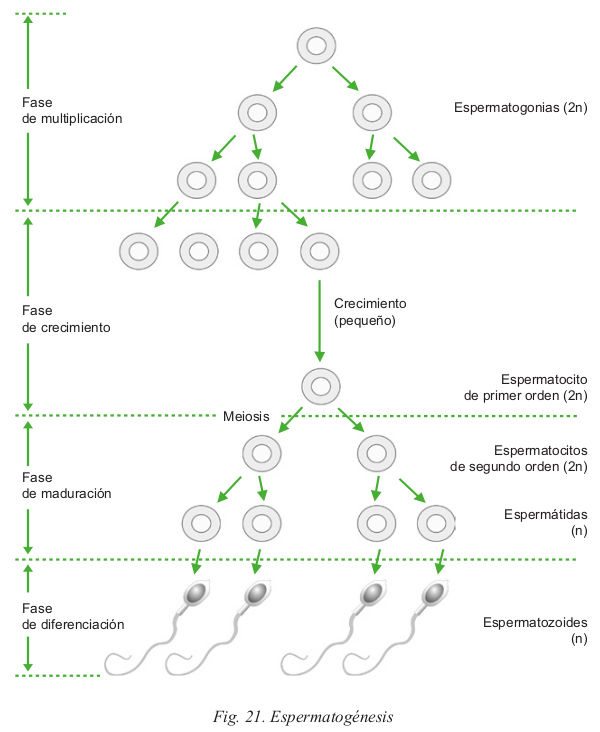
Comparación entre ovogénesis y espermatogénesis
Se acumula mayor cantidad de material nutritivo durante la ovogénesis que en la espermatogénesis.
Las células resultantes de la ovogénesis presentan tamaños distintos debido a que el material nutritivo no se distribuye equitativamente; en cambio, en la espermatogénesis todas sus células resultantes son de igual tamaño.
En la ovogénesis se produce sólo un gameto funcional. Al contrario, en la espermatogénesis se producen cuatro.
En la espermatogénesis se requiere un proceso de diferenciación celular, espermiogénesis, para obtener gametos funcionales. En la ovogénesis no.
La ovogénesis se inicia en la mujer en el tercer mes del desarrollo intrauterino. En el hombre, la espermatogénesis, cuando éste alcanza la pubertad.
Los ovocitos primarios, de la ovogénesis, quedan retenidos en la premeiosis hasta el momento de la ovulación. Los espermatocitos primarios continúan su proceso de división meiótica.
El óvulo
El óvulo es el gameto femenino que se ha formado en las gónadas femeninas, los ovarios, a lo largo de la vida fértil de la mujer. Se pueden distinguir varias partes dentro de un óvulo (Fig.22):
- Pronúcleo ovular: el pronúcleo es el núcleo de los gametos. Posee la mitad del número de cromosomas, células haploides (n), de los núcleos de las células somáticas. Durante la fecundación, los pronúcleos de un óvulo y un espermatozoide se fusionan, creando así la primera célula diploide del nuevo ser, llamada cigoto o célula huevo.
- Membrana plasmática: la membrana plasmática o celular es una estructura laminar que engloba a las células y define sus límites. Además, se asemeja a las membranas que delimitan los orgánulos de las células. Está compuesta por una lámina que sirve de “contenedor” para el citosol y los distintos compartimentos internos de la célula, así como también otorga protección mecánica. Está formada principalmente por fosfolípidos (fosfatidiletanolamina y fosfatidilcolina), colesterol, glúcidos y proteínas (integrales y periféricas). La principal característica de esta barrera es su permeabilidad selectiva, lo que le permite seleccionar las moléculas que deben entrar y salir de la célula. De esta manera se mantiene estable el medio intracelular, regulando el paso de agua, iones y metabolitos, a la vez que mantiene el potencial electroquímico (haciendo que el medio interno esté cargado negativamente). Cuando una molécula de gran tamaño atraviesa o es expulsada de la célula y se invagina parte de la membrana plasmática para recubrirlas cuando están en el interior ocurren respectivamente los procesos de endocitosis y exocitosis. Esta estructura celular contribuye a mantener el equilibrio entre el interior (medio intracelular) y el exterior (medio extracelular) de éstas.
- Espacio perivitelino: espacio que queda entre el ovocito y la zona pelúcida que lo envuelve, de un grosor aproximado de entre 0,2 y 0,4 micras.
- Membrana vitelina (Zona Pelúcida): se encuentra por fuera de la membrana citoplasmática. En mamíferos, incluido el ser humano, es muy gruesa y se denomina Zona Pelúcida (ZP). Se define, pues, como la capa externa que rodea el ovocito de los mamíferos en el folículo de Graaf, separándolo del espacio perivitelino. Está compuesta por varias glicoproteínas agrupadas en tres familias: ZP1, ZP2 y ZP3, según sus propiedades inmunológicas y funcionales. La zona pelúcida se encarga de la protección del ovocito y preembrión en sus primeros días de desarrollo, confinándolo en un volumen pequeño. Nuevas investigaciones llevadas a cabo demuestran que las glicoproteínas que la forman poseen receptores para los espermatozoides facilitando la fecundación. También tienen un papel en la reacción acrosómica induciendo la misma. El endurecimiento posterior a la entrada del espermatozoide debido a una segunda despolarización de la membrana (provocada por iones de calcio) es fundamental para el bloqueo poliespermático.
- Gránulos corticales: los gránulos corticales son orgánulos esféricos de pequeño tamaño (entre 100 y 200 nm de diámetro) que están presentes en el citoplasma de los gametos femeninos. Están delimitados por una membrana simple, y en su interior se almacenan sustancias de carácter glicoproteico, entre las que se encuentran fundamentalmente enzimas hidrolíticos. Tiene una importante función en la prevención de fecundaciones polispérmicas. Una vez que el ovocito ha sido penetrado por un espermatozoide, se produce la reacción cortical, que consiste en la exocitosis de los gránulos corticales.
- Corona radiata: es la estructura que rodea a un óvulo no fertilizado. Consiste en dos o tres capas de células foliculares unidas a la capa protectora más externa del óvulo, la zona pelúcida. Su función principal es proveer de proteínas a la célula huevo. La corona radiata aparece durante la ovulación, pero puede desaparecer después de la fertilización.

Espermatozoide
Es el gameto masculino que se produce de forma continua en las gónadas masculinas, los testículos, gracias a la interacción con las hormonas cuando se alcanza la pubertad. Las partes de un espermatozoide son las siguientes (Fig. 23 a y b):
- Acrosoma: el acrosoma es un pequeño depósito situado en el extremo apical de la cabeza del espermatozoide y que contiene enzimas hidrolíticas. Está limitado por la membrana acrosomal externa (adosada a la membrana celular) y por la membrana acrosomal interna (adosada a la membrana nuclear). El acrosoma se forma durante el proceso de espermatogénesis, a partir de la fusión de vesículas procedentes del aparato de Golgi, orgánulo donde se forman las enzimas antes mencionadas. Su función es debilitar y romper las distintas paredes que envuelven al óvulo.
- Cabeza: la cabeza es la parte fecundadora; es la parte más importante del espermatozoide ya que contiene la carga genética (23 cromosomas, en el pronúcleo) que unidos a los 23 del óvulo dan lugar a la célula madre formando 46 cromosomas agrupados en pares. La cabeza contiene el núcleo (que contiene la información genética) y el acrosoma (que contiene sustancias hidrolíticas para la penetración en el óvulo). Su función es insertar la cabeza (del espermatozoide) en el óvulo, en la fecundación, y transmitir la carga genética contenida en el pronúcleo masculino, que, unidos a los 23 del óvulo dan lugar a la célula madre, (al sumarse el total de 46 cromosomas, agrupados en pares).
- Pieza intermedia: con mitocondrias, que proporcionan la energía que necesita el espermatozoide para mover el flagelo y poder desplazarse; y el comienzo del filamento axial (que se prolongará en la cola). Entre la cabeza y la pieza intermedia se encuentra el centriolo (que actúa como “motor” del flagelo).
- Mitocondrias: sirven para darle energía a las estructuras citológicas que componen el flagelo de espermatozoide y permitir su movimiento.
- Flagelo: en biología es un orgánulo filiforme cuya función principal (aunque no
única) suele ser proveer de movimiento a las células. Este orgánulo presenta dos
partes:
- Filamento axial: es el que permite el movimiento del espermatozoide.
- Vaina de la cola: es la que regula el movimiento de la cola del espermatozoide. Es la parte más larga del espermatozoide. Mediante sus movimientos hace que el espermatozoide nade con la cabeza en primer término. Rodeado por una vaina proteica con 9 filamentos externos y dos centrales.
El constituyente fundamental de estos filamentos es la tubulina, proteína contráctil similar a las proteínas contráctiles del músculo, la actina y miosina.
La tubulina espermática se contrae en presencia de ATP y esta presencia está asociada a la enzima dineína que rompe las moléculas de ATP.